ดาวน์โหลดงานนำเสนอ
งานนำเสนอกำลังจะดาวน์โหลด โปรดรอ

1
ฮอร์โมน รองศาสตราจารย์ ดร. ดนัย บุณยเกียรติ
รองศาสตราจารย์ ดร. ดนัย บุณยเกียรติ ภาควิชาพืชสวน คณะเกษตรศาสตร์ มหาวิทยาลัยเชียงใหม่

2
ฮอร์โมน คือ สารประกอบอินทรีย์ ซึ่งสามารถมีผลกระทบในปริมาณที่น้อยมาก โดยพืชจะสังเคราะห์ที่ส่วนหนึ่งแล้วเคลื่อนย้ายไปยังอีกส่วนหนึ่ง และมีผลต่อกระบวนการทางสรีรวิทยาที่ค่อนข้างเฉพาะเจาะจง

3
สารควบคุมการเจริญเติบโต (Plant Growth Regulator)เป็นสารเคมีที่สำคัญในการเกษตรเป็นสารอินทรีย์ซึ่งมนุษย์สังเคราะห์ขึ้นมาได้ ซึ่งบางชนิดมีคุณสมบัติเหมือนฮอร์โมนพืช ในบางกรณีนี้ฮอร์โมนจะทดแทนสภาพแวดล้อมที่เหมาะสมได้ ในประเทศไทยการใช้ฮอร์โมนพืชมีวัตถุประสงค์ในทางการเกษตรเพื่อให้มีผลผลิต เพื่อเพิ่มผลผลิตและคุณภาพ และเพื่อความสะดวกในการจัดการฟาร์ม

4
ออกซิน (Auxin) Charles Darwin ซึ่งศึกษาเรื่อง Phototropism ซึ่งพืชจะโค้งงอเข้าหาแสง Darwin ทดลองกับ ต้นกล้าของ Phalaris canariensis และพบว่าโคลีออพไทล์ของพืชชนิดนี้จะตอบสนองต่อการได้รับแสงเพียงด้านเดียวทำให้เกิดการโค้งเข้าหาแสง Darwin สรุปว่าเมื่อต้นกล้าได้รับแสงจะทำให้มี "อิทธิพล" (Influence) บางอย่างส่งผ่านจากส่วนยอดมายังส่วนล่างของโคลีออพไทล์ ทำให้เกิดการโค้งงอเข้าหาแสง ซึ่งนักวิทยาศาสตร์รุ่นต่อมาได้ศึกษาถึง "อิทธิพล" ดังกล่าว
 บางอย่างส่งผ่านจากส่วนยอดมายังส่วนล่างของโคลีออพไทล์ ทำให้เกิดการโค้งงอเข้าหาแสง ซึ่งนักวิทยาศาสตร์รุ่นต่อมาได้ศึกษาถึง อิทธิพล ดังกล่าว.")
5
ออกซิน (Auxin) ต่อมา Boysen-Jensen และ Paal ได้ศึกษาและแสดงให้เห็นว่า "อิทธิพล" ดังกล่าวนี้มีสภาพเป็นสารเคมี Went ได้ทำงานทดลองและสามารถแยกสารชนิดนี้ออกจากโคลีออพไทล์ได้ โดยตัดส่วนยอดของโคลีออพไทล์ของข้าวโอ๊ตแล้ววางลงบนวุ้นจะทำให้สารเคมีที่กระตุ้นการเจริญเติบโตไหลลงสู่วุ้น เมื่อนำวุ้นไปวางลงที่ด้านหนึ่งของโคลีออพไทล์ที่ไม่มียอดด้านใดด้านหนึ่งจะทำให้โคลีออพไทล์ดังกล่าวโค้งได้

6
ออกซิน (Auxin) สารเคมีดังกล่าวได้รับการตั้งชื่อว่า ออกซิน ซึ่งในปัจจุบันพบในพืชชั้นสูงทั่วๆ ไป และมีความสำคัญต่อการเจริญเติบโตของพืช สังเคราะห์ได้จากส่วนเนื้อเยื่อเจริญของลำต้น ปลายราก ใบอ่อน ดอกและผล และพบมากที่บริเวณเนื้อเยื่อเจริญโคลีออพไทด์และคัพภะ รวมทั้งใบที่กำลังเจริญด้วย

7
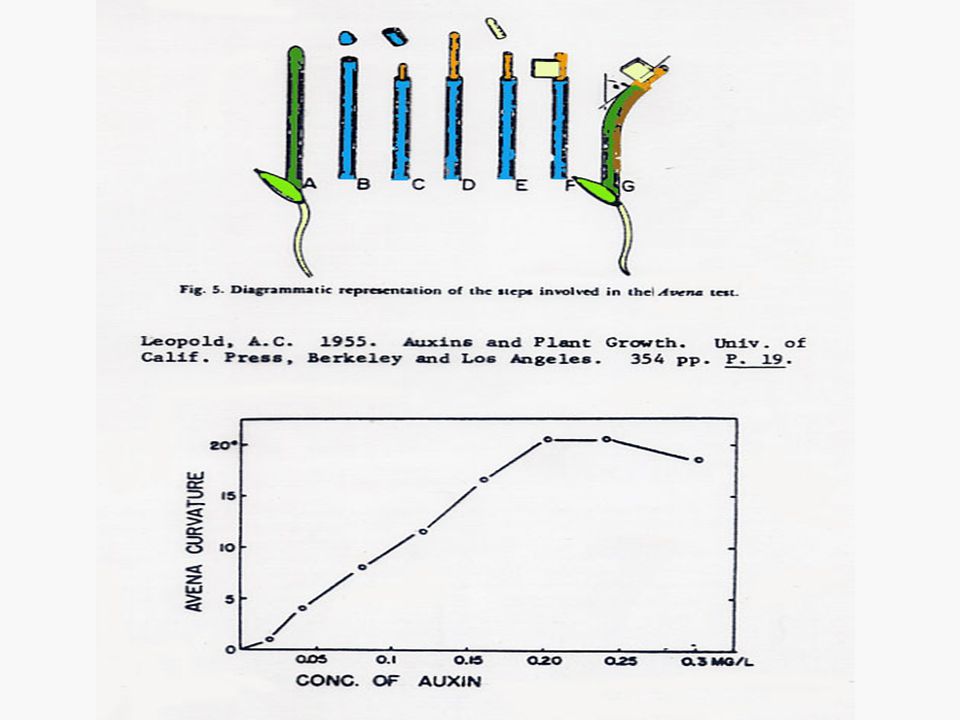
งานทดลองของนักวิทยาศาสตร์ ซึ่งนำมาสู่การค้นพบออกซินในพืช ทุกการทดลอง ทำกับโคลีออพไทด์ของต้นกล้าของพืชตระกูลหญ้า ลูกศร 3 เส้นที่ติดกัน แสดงทิศทางของ unilateral light ในการทดลองของ Went แสดงถึง Went Avena curvature test

8
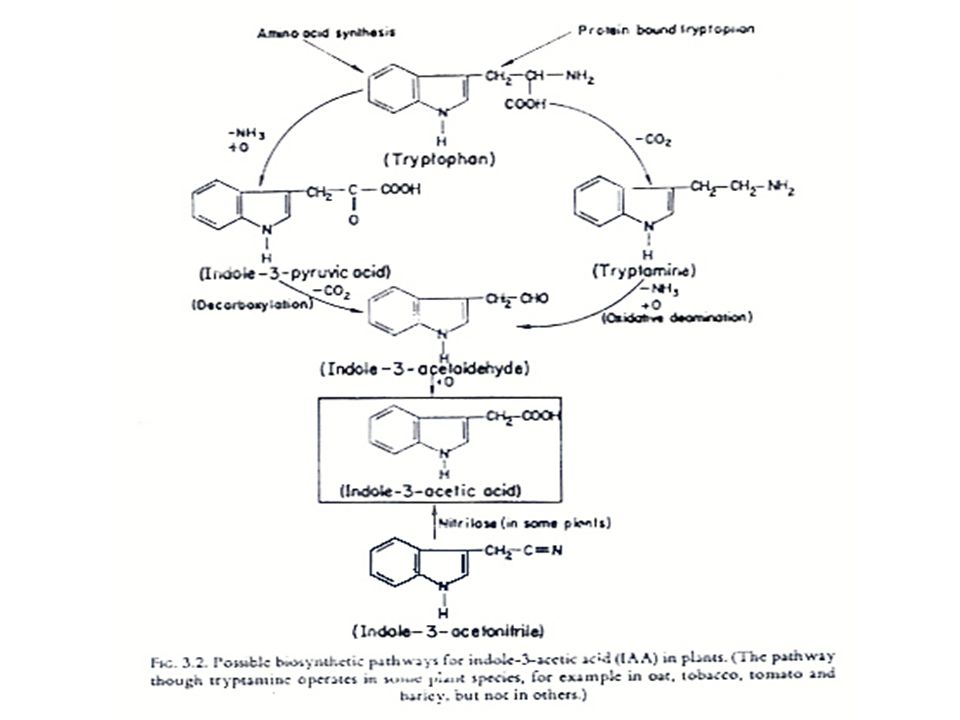
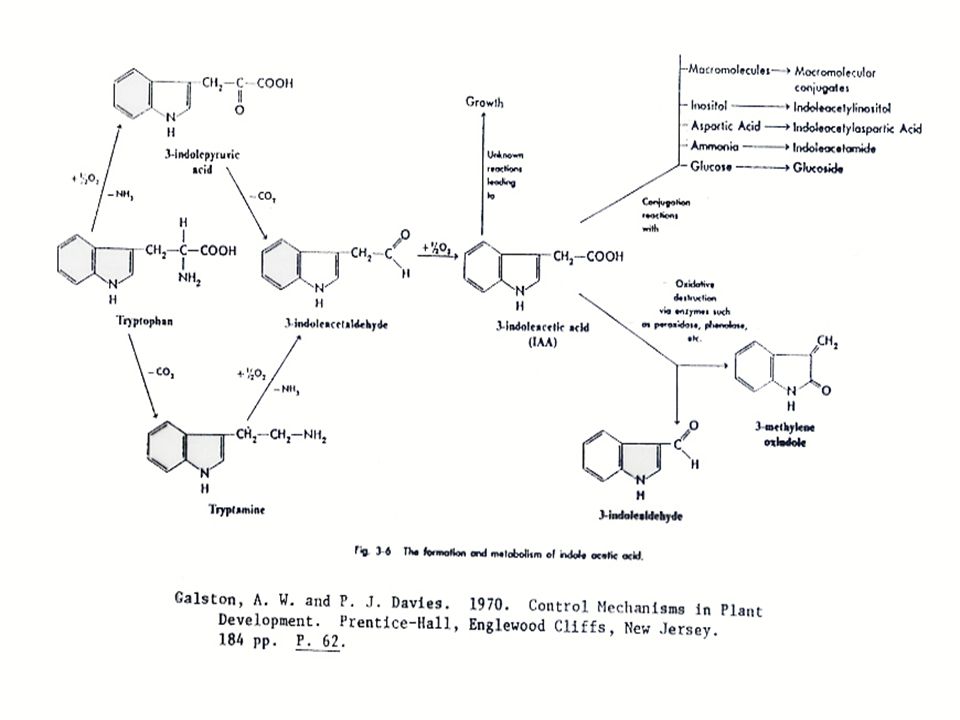
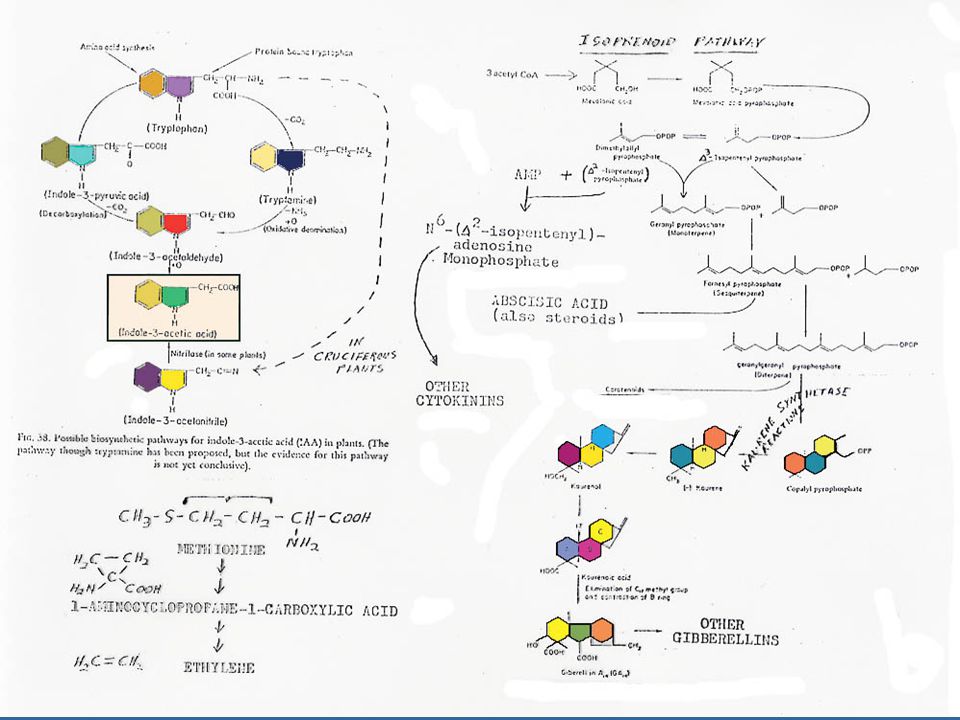
การสังเคราะห์ออกซิน การสังเคราะห์ออกซินนั้นมีกรดอะมิโน L-Tryptophan เป็นสารเริ่มต้น (Precursor) L-Tryptophan เป็นกรดอะมิโนที่มีโครงสร้างของ Indole อยู่ การสังเคราะห์ออกซิน แสดงในรูปที่ 12.2 ซึ่งในการสังเคราะห์ IAA นั้น จะมี IAAld และ IPyA เป็นสารที่พบในระหว่างการสังเคราะห์ ในพืชบางชนิด เช่น ข้าวโอ๊ต ยาสูบ มะเขือเทศ ทานตะวัน และข้าวบาร์เลย์ พบว่า Tryptophan สามารถเปลี่ยนเป็น Tryptamine ได้ ในพืชตระกูลกะหล่ำ Tryptamine อาจจะเปลี่ยนไปเป็น Indoleacetaldoxime แล้วเปลี่ยนไปเป็น IAN แล้วจึงเปลี่ยนเป็น IA
 L-Tryptophan เป็นกรดอะมิโนที่มีโครงสร้างของ Indole อยู่ การสังเคราะห์ออกซิน แสดงในรูปที่ 12.2 ซึ่งในการสังเคราะห์ IAA นั้น จะมี IAAld และ IPyA เป็นสารที่พบในระหว่างการสังเคราะห์ ในพืชบางชนิด เช่น ข้าวโอ๊ต ยาสูบ มะเขือเทศ ทานตะวัน และข้าวบาร์เลย์ พบว่า Tryptophan สามารถเปลี่ยนเป็น Tryptamine ได้ ในพืชตระกูลกะหล่ำ Tryptamine อาจจะเปลี่ยนไปเป็น Indoleacetaldoxime แล้วเปลี่ยนไปเป็น IAN แล้วจึงเปลี่ยนเป็น IA.")
10
การสังเคราะห์ออกซิน การศึกษาเรื่องการสังเคราะห์ออกซินมักศึกษาจากเนื้อเยื่อปลายรากหรือปลายยอด และพบว่า IAA นี้สังเคราะห์ได้ทั้งในส่วนไซโตซอล (Cytosol) ไมโต-คอนเดรีย และคลอโรพลาสต์ ในการศึกษาในปัจจุบันพบว่า Phenylacetic acid หรือ PAA มีคุณสมบัติของออกซินด้วยและสามารถสังเคราะห์ได้จาก L-Phenylalanine โดยพบในคลอโรพลาสต์ และไมโตคอนเดรียของทานตะวัน
 ไมโต-คอนเดรีย และคลอโรพลาสต์ ในการศึกษาในปัจจุบันพบว่า Phenylacetic acid หรือ PAA มีคุณสมบัติของออกซินด้วยและสามารถสังเคราะห์ได้จาก L-Phenylalanine โดยพบในคลอโรพลาสต์ และไมโตคอนเดรียของทานตะวัน.")
11
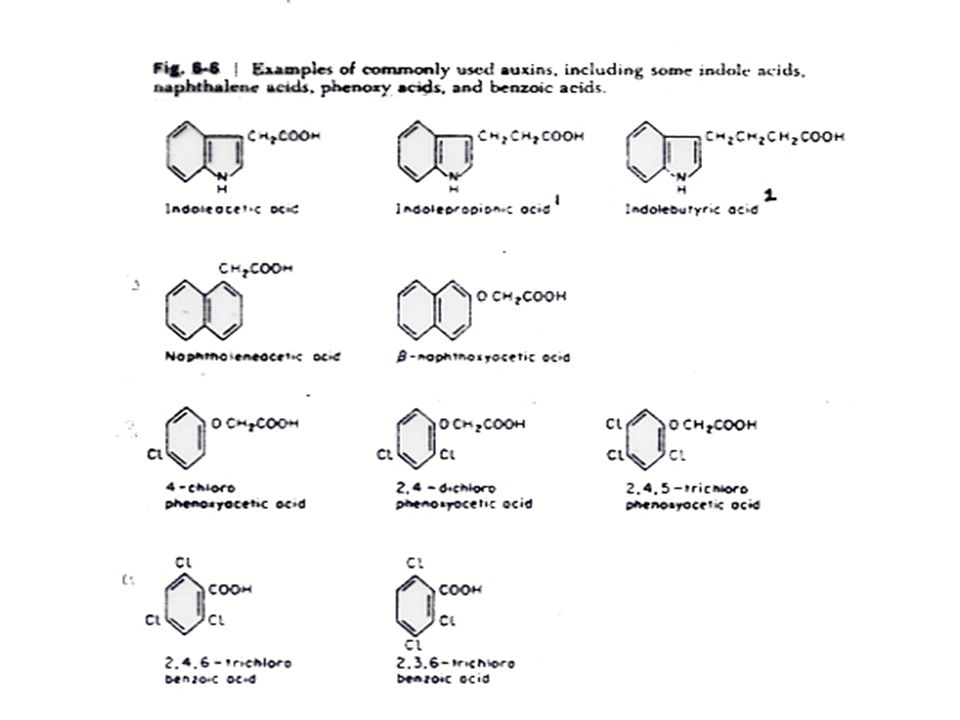
การสังเคราะห์ออกซิน สารสังเคราะห์ที่มีคุณสมบัติของออกซินมีหลายชนิดที่สำคัญทางการเกษตร เช่น สาร 2, 4-dichlorophenoxyacetic acid หรือ 2,4-D ซึ่งใช้ในการกำจัดวัชพืช IBA หรือ Indole butyric acid ใช้ในการเร่งให้ส่วนที่จะนำไปปักชำเกิดรากเร็วขึ้น และ NAA หรือ Napthalene acetic acid จะช่วยในการติดผลของผลไม้บางชนิด

13
โครงสร้างของโมเลกุลที่สำคัญของสารที่จะมีคุณสมบัติของออกซินคือ ต้องประกอบด้วยประจุลบ (Strong Negative Charge) ซึ่งเกิดจากการแตกตัวของกลุ่มคาร์บอกซิล และประจุลบจะต้องอยู่ห่างจากประจุบวก (Weaker Positive Charge) บนวงแหวนด้วยระยะทางประมาณ 5.5 Angstrom
 ซึ่งเกิดจากการแตกตัวของกลุ่มคาร์บอกซิล และประจุลบจะต้องอยู่ห่างจากประจุบวก (Weaker Positive Charge) บนวงแหวนด้วยระยะทางประมาณ 5.5 Angstrom")
14
การสลายตัวของ IAA 1. Photo-oxidation IAA ที่อยู่ในสภาพสารละลายจะสลายตัวได้เมื่อได้รับแสง การเกิด Photo-oxidation ของ IAA จะถูกเร่งโดยการปรากฏของสารสีตามธรรมชาติ หรือที่สังเคราะห์ได้ จึงอาจจะเป็นไปได้ว่าการที่สารสีของพืชดูดซับพลังงานจากแสงแล้วทำให้เกิดการออกซิไดซ์ IAA ซึ่งสารสีที่เกี่ยวข้อง คือ ไรโบฟลา-วิน และไวโอลาแซนธิน (Riboflavin และ Violaxanthin) สารที่เกิดขึ้นเมื่อ IAA สลายตัวโดยแสงคือ 3-methylene-2-oxindole และ Indoleacetaldehyde
 สารที่เกิดขึ้นเมื่อ IAA สลายตัวโดยแสงคือ 3-methylene-2-oxindole และ Indoleacetaldehyde.")
15
2. การออกซิไดซ์โดยเอนไซม์ (Enzymic Oxidation of IAA) พืชหลายชนิดมีเอนไซม์เรียกว่า IAA-oxidase ซึ่งจะคะตะไลท์ สลาย IAA ได้คาร์บอนไดออกไซด์ และเป็นปฏิกิริยาที่ใช้ออกซิเจน IAA-oxidase ผลิตภัณฑ์ที่ได้ คือ 3-methylene-2-oxindole และถูกเมตาโบไลซ์ต่อไป เป็น 3-methyl-2-oxindole
 พืชหลายชนิดมีเอนไซม์เรียกว่า IAA-oxidase ซึ่งจะคะตะไลท์ สลาย IAA ได้คาร์บอนไดออกไซด์ และเป็นปฏิกิริยาที่ใช้ออกซิเจน IAA-oxidase ผลิตภัณฑ์ที่ได้ คือ 3-methylene-2-oxindole และถูกเมตาโบไลซ์ต่อไป เป็น 3-methyl-2-oxindole.")
16
3. รวมกับสารชนิดอื่นในไซโตพลาสต์
4. เปลี่ยนเป็นอนุพันธ์ชนิดอื่น

17
1. Bioassay คือ การวัดปริมาณออกซินโดยใช้ชิ้นส่วนของพืช เช่น โคลีออพไทล์ของข้าวโอ๊ตหรือพืชใบเลี้ยงเดี่ยวอื่นๆ แล้ววัดความโค้งของยอดโดยการวางออกซินที่ต้องการวัดปริมาณลงบนส่วนของโคลีออพไทล์ซึ่งตัดยอดออกแล้ว มุมที่โค้งจะบอกปริมาณของออกซินได้โดยเปรียบเทียบจากเส้นมาตรฐาน (Standard Curve)
.")
21
2. การวัดจากคุณสมบัติทางฟิสิกส์ สกัดจนเป็นสารบริสุทธิ์แล้วจึงใช้เครื่อง Gas Chromatograph ร่วมกับ Mass Spectrometry ในการจำแนกและหาปริมาณ 3. การวัดโดยวิธีเคมีโดยให้ออกซินทำปฏิกิริยากับ Salkowski's Reagent (acidified ferric chloride) หรือใช้ Ehrllch's Reagent ซึ่งจะเกิดสีขึ้นมา จากนั้นวัดความเข้มของสีแล้วเปรียบเทียบกับเส้นมาตรฐาน
 หรือใช้ Ehrllch s Reagent ซึ่งจะเกิดสีขึ้นมา จากนั้นวัดความเข้มของสีแล้วเปรียบเทียบกับเส้นมาตรฐาน.")
22
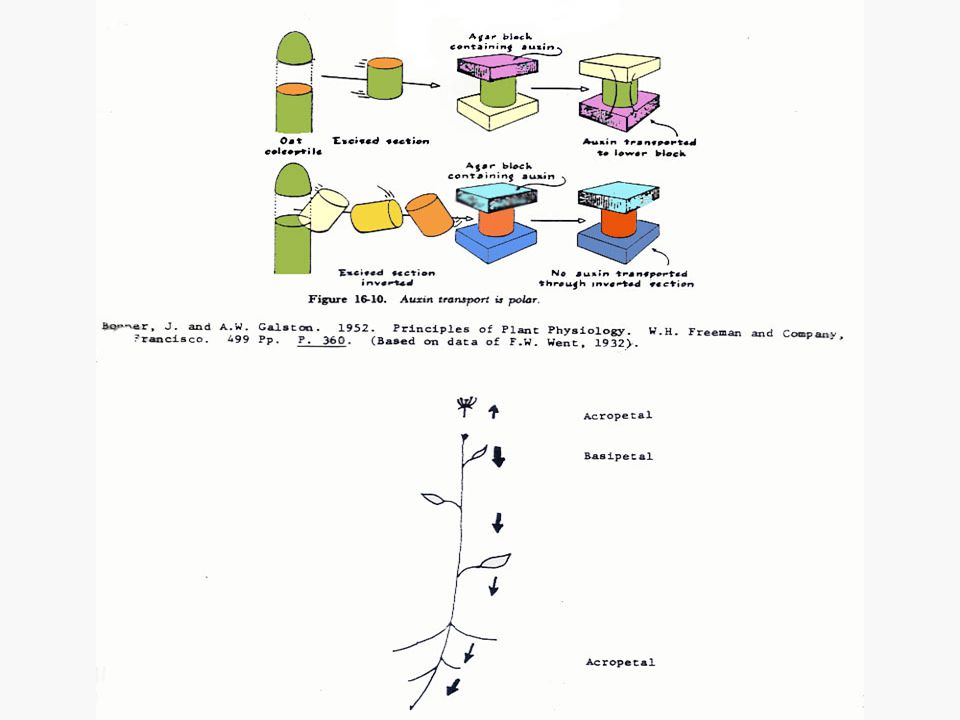
การเคลื่อนที่ของออกซินในต้นพืช
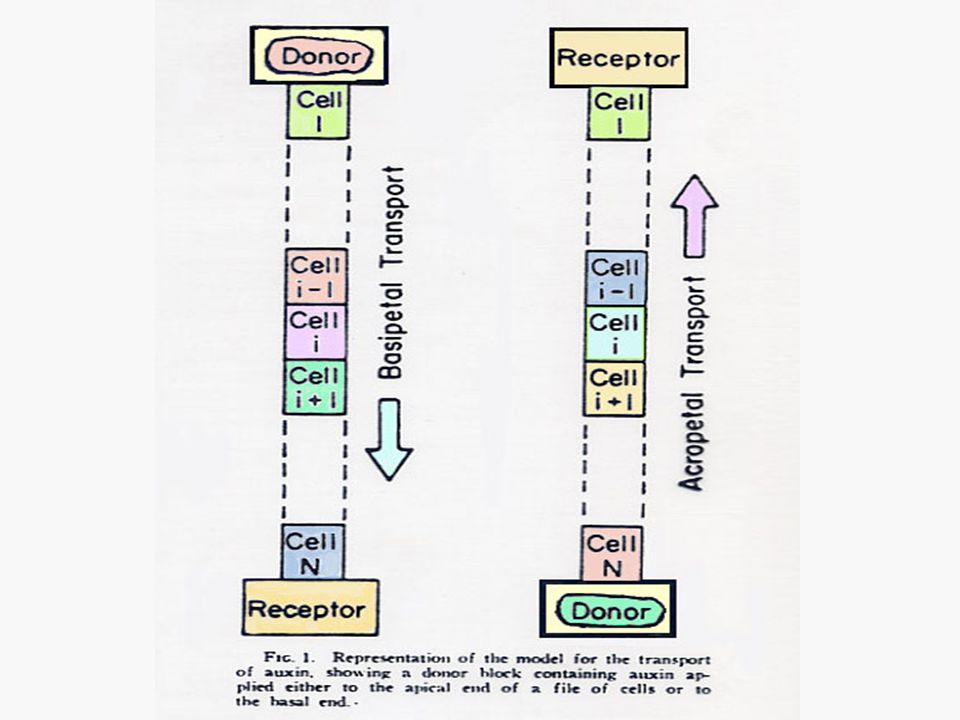
การเคลื่อนที่ของออกซินจะเป็นแบบโพลาไรซ์ (Polarized) คือ เคลื่อนที่ไปตามยาวของลำต้นโดยไปในทิศทางใดทิศทางหนึ่งมากกว่าทิศทางตรงกันข้าม การเคลื่อนที่ของออกซินในส่วนที่อยู่เหนือดิน จะเป็นแบบโพลาร์ เบสิพีตัล (Polar Basipetal) คือ จะเคลื่อนที่จากแหล่งผลิตที่ยอดไปสู่โคนต้น การเคลื่อนที่ของออกซินจะเกิดแบบเบสิพีตัลก็เมื่อท่อนเนื้อเยื่อวางอยู่ในลักษณะปกติของลักษณะทางสัณฐานวิทยาเท่านั้น
 คือ เคลื่อนที่ไปตามยาวของลำต้นโดยไปในทิศทางใดทิศทางหนึ่งมากกว่าทิศทางตรงกันข้าม. การเคลื่อนที่ของออกซินในส่วนที่อยู่เหนือดิน จะเป็นแบบโพลาร์ เบสิพีตัล (Polar Basipetal) คือ จะเคลื่อนที่จากแหล่งผลิตที่ยอดไปสู่โคนต้น การเคลื่อนที่ของออกซินจะเกิดแบบเบสิพีตัลก็เมื่อท่อนเนื้อเยื่อวางอยู่ในลักษณะปกติของลักษณะทางสัณฐานวิทยาเท่านั้น.")
30
ในกรณีของโคลีออพไทล์ของพืชนั้นชี้ให้เห็นว่าออกซินเคลื่อนที่ผ่านเซลล์ทุกเซลล์ลงมาแต่ในกรณีของลำต้นนั้นยังไม่มีหลักฐานชี้ให้เห็นเด่นชัดนัก แต่อาจจะเป็นไปได้ว่าโปรแคมเบียม (Procambium) และแคมเบียม (Cambium) การเคลื่อนที่ของออกซินในรากก็มีลักษณะเป็นโพลาร์ แต่เป็นแบบ อะโครพีตัล

31
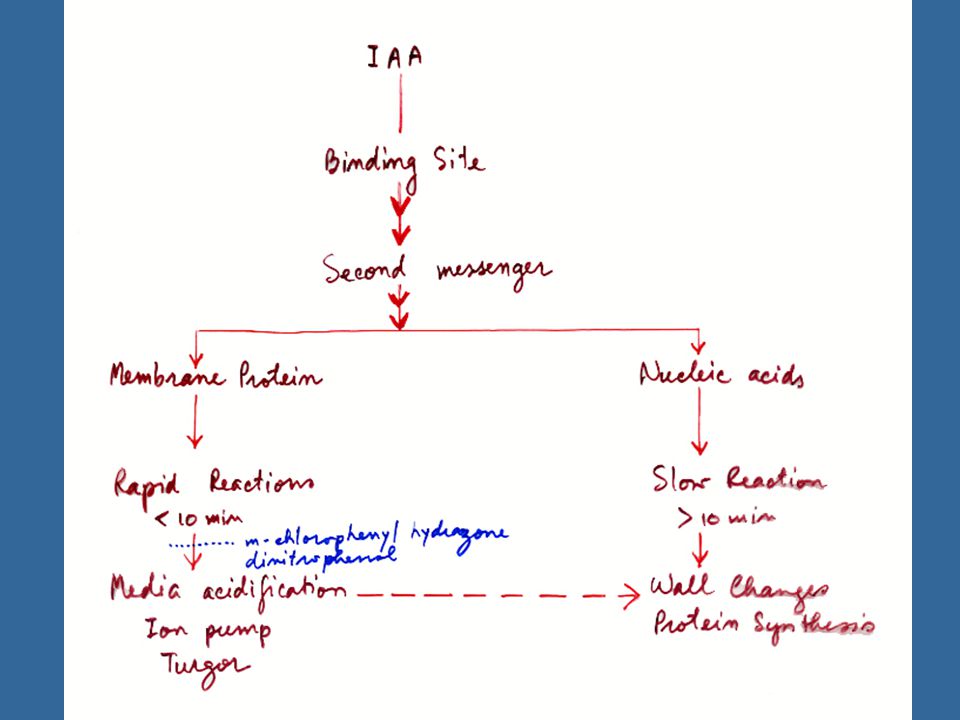
กลไกในการทำงานของออกซินในระยะที่ผ่านมาจะมีแนวความคิดเป็นสองอย่าง คือ แนวคิดที่เกี่ยวข้องกับผนังเซลล์เป็นส่วนที่รับผลกระทบของออกซิน และขยายตัว ส่วนอีกแนวคิดหนึ่งมุ่งไปที่ผลของออกซินต่อเมตาบอลิสม์ของกรดนิวคลีอิคในปัจจุบันได้นำสองแนวคิดมาวิเคราะห์ ร่วมกันเพื่อศึกษากลไกในการทำงานของออกซิน และยังศึกษาผลของออกซินต่อเยื่อหุ้มเซลล์ด้วย

32
การขยายตัวของเซลล์จะสัมพันธ์กับการเปลี่ยนแปลงปริมาณและกิจกรรมของเอนไซม์ โดยที่ออกซินจะมีบทบาทต่อ กระบวนการเมตาบอลิสม์ของกรดนิวคลีอิค โดยการศึกษาจากการเพาะเลี้ยงเนื้อเยื่อที่เป็นไส้ของต้นยาสูบ (Tobacco Pith) ซึ่งจะเจริญไปเป็นกลุ่มเนื้อเยื่อ (Callus) นั้นพบว่ามีปริมาณของ RNA เพิ่มมากขึ้น ทั้งนี้เพราะออกซินจะกระตุ้นให้มีการสังเคราะห์ RNA มากขึ้น แล้วส่งผลไปถึงการเจริญของกลุ่มเนื้อเยื่อ ถ้าหากใช้สารระงับการสังเคราะห์โปรตีนหรือ RNA ความสามารถในการกระตุ้นการเจริญเติบโตของออกซินจะหายไป
 ซึ่งจะเจริญไปเป็นกลุ่มเนื้อเยื่อ (Callus) นั้นพบว่ามีปริมาณของ RNA เพิ่มมากขึ้น ทั้งนี้เพราะออกซินจะกระตุ้นให้มีการสังเคราะห์ RNA มากขึ้น แล้วส่งผลไปถึงการเจริญของกลุ่มเนื้อเยื่อ ถ้าหากใช้สารระงับการสังเคราะห์โปรตีนหรือ RNA ความสามารถในการกระตุ้นการเจริญเติบโตของออกซินจะหายไป")
33
ออกซินมีผลต่อระดับเอนไซม์ โดยผ่านทางการสังเคราะห์ RNA นอกจากนั้นออกซินยังมีผลกระทบต่อกิจกรรมของเอนไซม์โดยตรง เช่น การกระตุ้นให้เอนไซม์เกิดกิจกรรมหรือเปลี่ยนรูปมาอยู่ในรูปที่มีกิจกรรมได้ นักวิทยาศาสตร์ได้มุ่งความสนใจไปสู่เอนไซม์ที่สัมพันธ์กับกระบวนการขยายตัวของเซลล์ เซลล์พืชจะมีผนังเซลล์อยู่ข้างนอกสุด ดังนั้นการเจริญของเซลล์จะเกิดขึ้นได้เมื่อคุณสมบัติของผนังเซลล์เปลี่ยนไปในทางที่ก่อให้เกิดการขยายตัวของโปรโตพลาสต์

34
เซลล์พืชทุกชนิดที่ผ่านขั้นตอนของเนื้อเยื่อเจริญมาแล้วจะผ่านขั้นตอนการเจริญเติบโต 2 ขั้น คือ การแบ่งเซลล์และการขยายตัวแวคคิวโอขึ้นภายในเซลล์ (Vacuolation) การขยายตัวของเซลล์นั้น ไม่เพียงแต่ผนังเซลล์ยืดตัวเท่านั้น แต่ยังมีการเพิ่มความหนาของผนังเซลล์เพราะมีสารใหม่ ๆ ไปเกาะด้วย

35
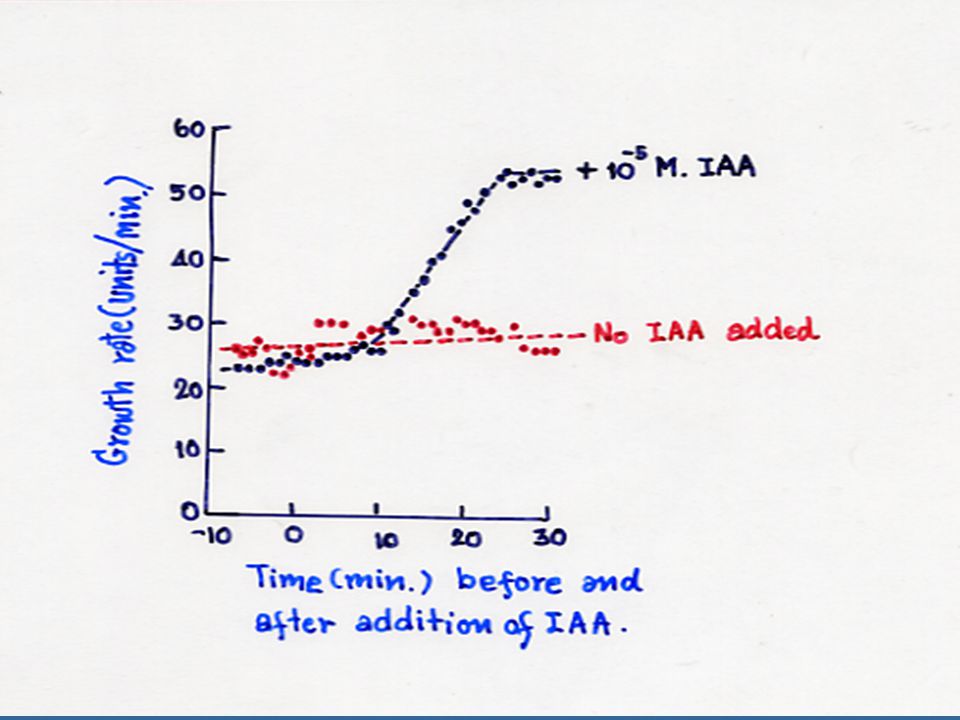
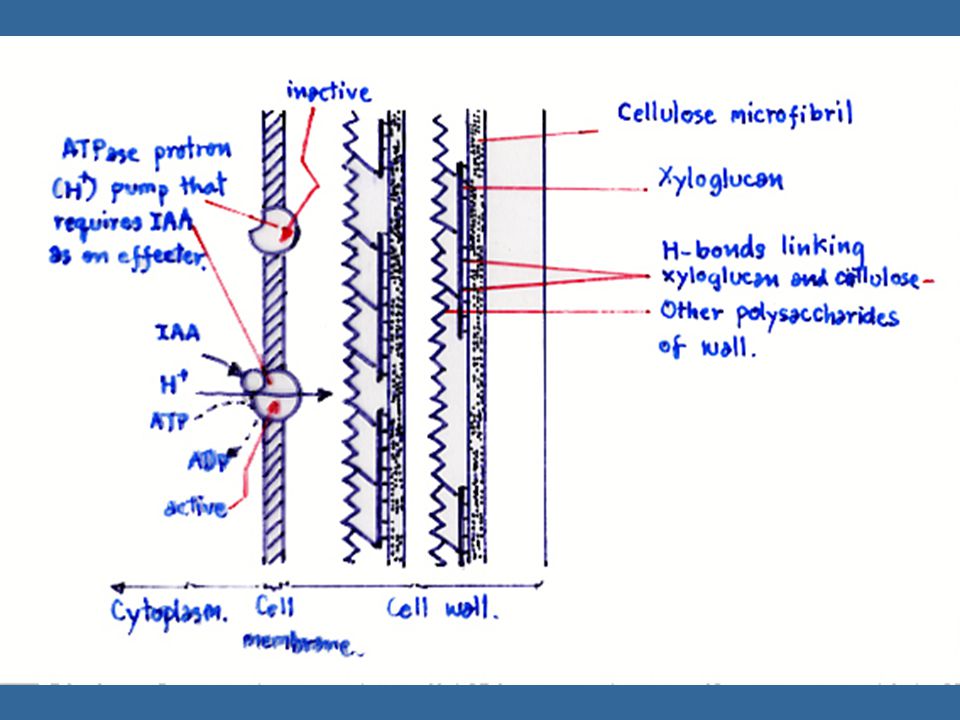
นักวิทยาศาสตร์ได้นำเอาโคลีออพไทล์หรือลำต้นที่ไม่ได้รับแสงไปแช่ลงในสภาพที่มี pH ประมาณ 3 ปรากฏว่าโคลีออพไทล์และลำต้นสามารถยืดตัวได้ และเรียกปรากฏการณ์นี้ว่า "Acid Growth Effect" ซึ่งให้ผลเหมือนกับการให้ออกซินแก่พืช การทดลองนี้ได้นำไปสู่การศึกษาที่แสดงว่าออกซิน กระตุ้นการปลดปล่อย H+ หรือโปรตอนจากเนื้อเยื่อ ทำให้ pH ของผนังเซลล์ต่ำลง ซึ่งการปลดปล่อย H+ นี้ต้องใช้พลังงานจากการหายใจด้วย สมมุติฐานเกี่ยวกับ "Proton-Pump" นี้ คาดว่าเกิดในเยื่อหุ้มเซลล์

36
การเจริญของเซลล์ต้องการ RNA และโปรตีนในช่วงที่เซลล์ยืดตัว เพราะในการยืดตัวของเซลล์นั้นผนังเซลล์ไม่ได้บางลงไป แต่ยังคงหนาเท่าเดิมหรือหนาขึ้น ดังนั้นจึงต้องมีการสร้างผนังเซลล์เพิ่มขึ้นด้วย ในการสร้างผนังเซลล์นั้นต้องใช้เอนไซม์และ RNA pH ต่ำมีผลต่อการเปลี่ยนคุณสมบัติของผนังเซลล์ในแง่ที่ว่า แขนที่เกาะกันของผนังเซลล์นั้นอาจจะถูกทำลายในสภาพที่ pH ต่ำ หรืออาจจะเป็น pH ที่เหมาะสมสำหรับเอนไซม์ ที่จะทำให้ผนังเซลล์เปลี่ยนไป

40
การตอบสนองของพืชต่อออกซิน
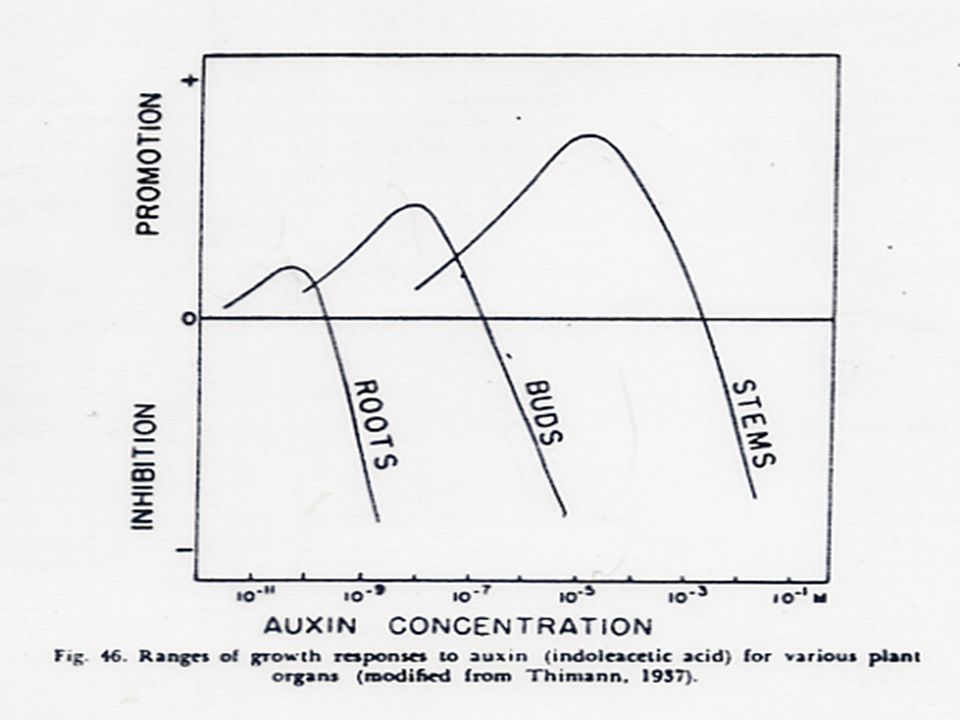
1. การตอบสนองในระดับเซลล์ออกซินทำให้เกิด การขยายตัวของเซลล์ (Cell enlargement) 2. การตอบสนองของอวัยวะหรือพืชทั้งต้น 2.1 เกี่ยวข้องกับการตอบสนองของพืชต่อแสง (Phototropism) Geotropism ดังได้กล่าวมาแล้ว 2.2 การที่ตายอดข่มไม่ให้ตาข้างเจริญเติบโต (Apical Dominance)
 2. การตอบสนองของอวัยวะหรือพืชทั้งต้น. 2.1 เกี่ยวข้องกับการตอบสนองของพืชต่อแสง (Phototropism) Geotropism ดังได้กล่าวมาแล้ว. 2.2 การที่ตายอดข่มไม่ให้ตาข้างเจริญเติบโต (Apical Dominance)")
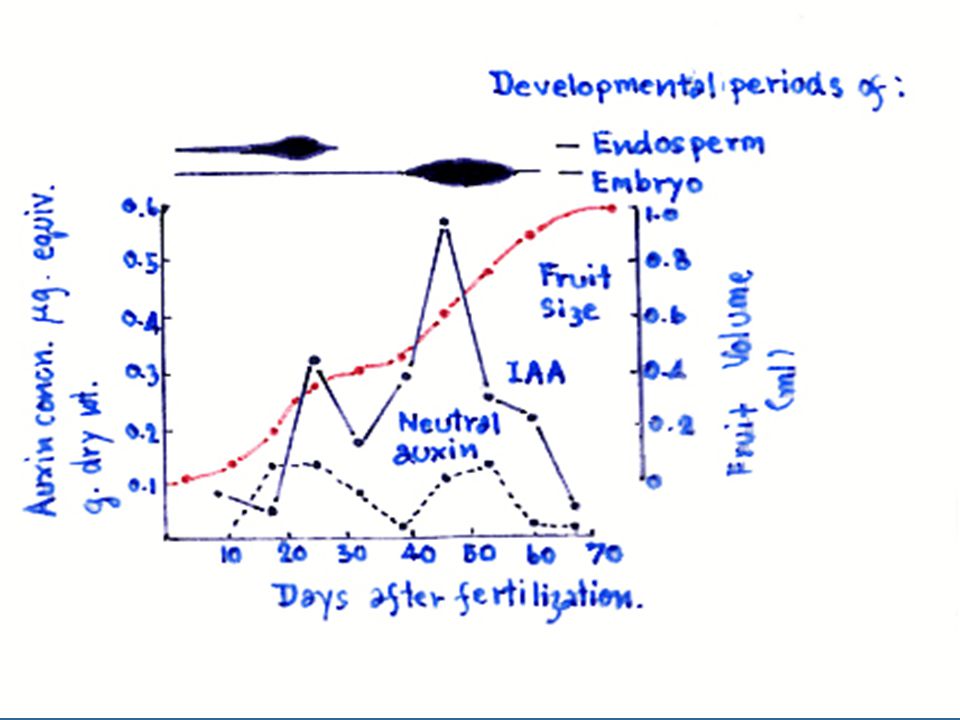
41
2.3 การติดผล เช่น กรณีของมะเขือเทศ ออกซินในรูปของ 4 CPA จะเร่งให้เกิดผลแบบ Pathenocarpic และในเงาะถ้าใช้ NAA 4.5 เปอร์เซ็นต์ จะเร่งการเจริญของเกสรตัวผู้ทำให้สามารถผสมกับเกสรตัวเมียได้ ในดอกที่ได้รับ NAA เกสรตัวเมียจะไม่เจริญเพราะได้รับ NAA ที่มีความเข้มข้นสูงเกินไป แต่เกสรตัวผู้ยังเจริญได้ ทำให้การติดผลเกิดมากขึ้น 2.4 ป้องกันการร่วงของผลโดยออกซินจะยับยั้งไม่ให้เกิด Abcission layer ขึ้นมา เช่น การใช้ 2,4-D ป้องกันผลส้มไม่ให้ร่วง หรือ NAA สามารถป้องกันการร่วงของผลมะม่วง 2.5 ป้องกันการร่วงของใบ 2.6 ในบางกรณีออกซินสามารถทำให้สัดส่วนของดอกตัวเมีย และตัวผู้เปลี่ยนไปโดยออกซินจะกระตุ้นให้มีดอกตัวเมียมากขึ้น

42
จิบเบอเรลลิน (Gibberellins)
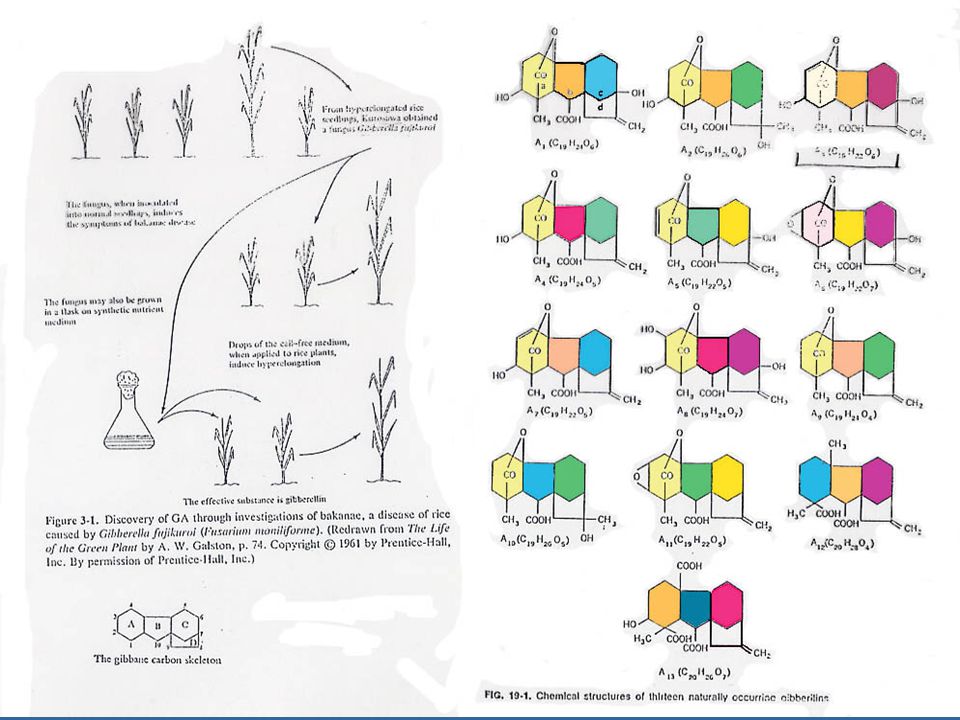
เมื่อ Kurosawa นักวิทยาศาสตร์ชาวญี่ปุ่น ศึกษาในต้นข้าวที่เป็นโรค Bakanae หรือโรคข้าวตัวผู้ ซึ่งเกิดจากเชื้อรา Gibberella fujikuroi หรือ Fusarium moniliforme ซึ่งทำให้ต้นข้าวมีลักษณะสูงกว่าต้นข้าวปกติ ทำให้ล้มง่าย ความรู้เกี่ยวกับโครงสร้างและส่วนประกอบทางเคมีของจิบเบอเรลลินนั้นได้รับการศึกษาในปี 1954 โดยนักเคมีชาวอังกฤษซึ่งสามารถแยกสารบริสุทธิ์จากอาหารเลี้ยงเชื้อรา Gibberella fujikuroi และเรียกสารนี้ว่ากรดจิบเบอเรลลิค (Gibberellic Acid)
")
44
ในปัจจุบันมีจิบเบอเรลลินซึ่งเป็นชื่อเรียกทั่ว ๆ ไปของสารประกอบประเภทนี้ประมาณไม่น้อยกว่า 80 ชนิด ชื่อเรียกสารประกอบชนิดนี้จะตั้งชื่อดังนี้ คือ Gibberellins A1 (GA1), A2, A3 เป็นต้น โดยที่กรดจิบเบอเรลลิค คือ GA3 GA ทุกชนิดจะมีโครงสร้างพื้นฐานของโมเลกุลเป็น Gibberellane Carbon Skeleton ซึ่งจะเหมือนกับกรดจิบเบอเรลลิค จะแตกต่างกันตรงจำนวนและตำแหน่งของกลุ่มที่เข้าแทนที่ในวงแหวนและระดับของความอิ่มตัวของวงแหวน A GA ประกอบด้วยคาร์บอนประมาณ 19-20 อะตอม ซึ่งจะรวมกันเป็นวงแหวน 4 หรือ 5 วงและจะต้องมีกลุ่มคาร์บอก-ซิล อย่างน้อย 1 กลุ่ม โดยใช้ชื่อย่อว่า GA

45
การสังเคราะห์จิบเบอเรลลิน
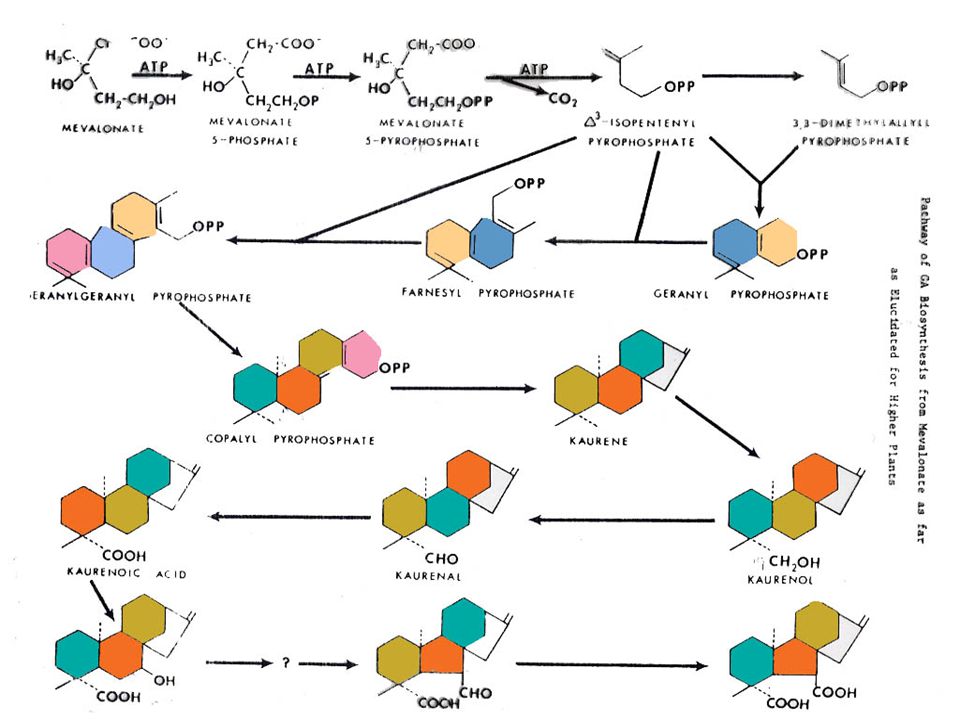
การสังเคราะห์ GA แสดงอยู่ในรูปที่ โดยที่สารเริ่มต้นเป็นกรดเมวาโลนิค (Mevalonic Acid) เปลี่ยนไปตามวิถีจนเกิดเป็นกรดคอรีโนอิค (Kaurenoic Acid) แล้วจึงเปลี่ยนไปเป็น GA ซึ่งวิถีในช่วงที่เปลี่ยนไปเป็น GA ชนิดต่าง ๆ นี้ยังไม่ทราบแน่ชัดนัก สารชนิดแรกที่มีวงแหวนของ Gibberellane คือ อัลดีไฮด์ของ GA12
 เปลี่ยนไปตามวิถีจนเกิดเป็นกรดคอรีโนอิค (Kaurenoic Acid) แล้วจึงเปลี่ยนไปเป็น GA ซึ่งวิถีในช่วงที่เปลี่ยนไปเป็น GA ชนิดต่าง ๆ นี้ยังไม่ทราบแน่ชัดนัก สารชนิดแรกที่มีวงแหวนของ Gibberellane คือ อัลดีไฮด์ของ GA12.")
46
การสังเคราะห์จิบเบอเรลลิน
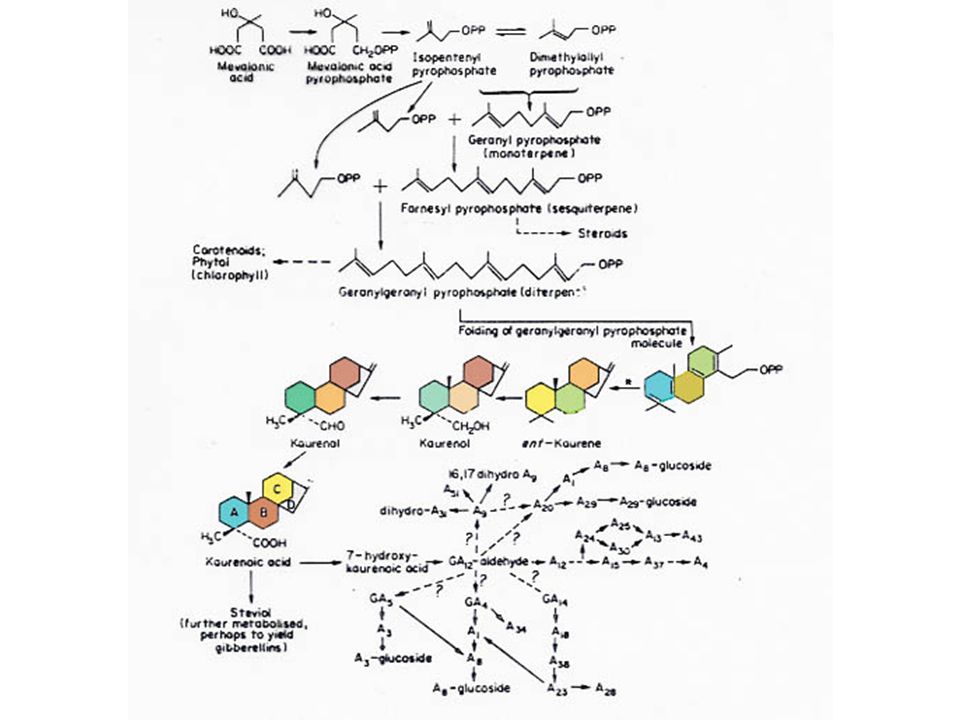
ในปัจจุบันมีสารชะงักการเจริญเติบโต เช่น CCC หรือ Cycocel AM Phosfon-D และ SADH หรือ Alar ซึ่งใช้กันมากในการเกษตร สารเหล่านี้บางชนิดสามารถระงับ กระบวนการสังเคราะห์จิบเบอเรลลินได้ เช่น AMO-1618 สามารถระงับการสังเคราะห์จิบเบอเรลลินในอาหารสำรอง (Endosperm) ของแตงกวาป่า โดยระงับในช่วงการเปลี่ยน Geranylgeranyl pyrophosphate ไปเป็น Kaurene ในทำนองเดียวกัน CCC สามารถระงับกระบวนการนี้ได้ด้วย
 ของแตงกวาป่า โดยระงับในช่วงการเปลี่ยน Geranylgeranyl pyrophosphate ไปเป็น Kaurene ในทำนองเดียวกัน CCC สามารถระงับกระบวนการนี้ได้ด้วย.")
51
การสังเคราะห์จิบเบอเรลลิน
ใบอ่อน ผลอ่อนและต้นอ่อนเป็นส่วนที่สร้าง GA ของพืช รากพืชอาจจะสามารถสร้าง GA ได้บ้าง GA ที่พบในปัจจุบันจึงเป็นสารธรรมชาติทั้งสิ้น

52
การสลายตัวของจิบเบอเรลลิน
จิบเบอเรลลินมีกิจกรรมทางสรีรวิทยาอยู่ได้เป็นระยะเวลานานในเนื้อเยื่อพืช จิบเบอเรลลินสามารถเปลี่ยนจากชนิดหนึ่งไปเป็นจิบเบอเรลลินอีกชนิดหนึ่งได้ในเนื้อเยื่อพืช จิบเบอเรลลินในรูปของไกลโคไซด์ (Glycosides) กรดจิบเบอเรลลิค ซึ่งอยู่ในสภาพสารละลายถูกทำให้สลายตัวได้โดยใช้ Acid Hydrolysis ที่อุณหภูมิสูงและได้ผลิตภัณฑ์คือกรดจิบเบอเรลลี-นิค (Gibberellenic Acid) และกรดจิบเบอริค (Gibberic Acid)
 กรดจิบเบอเรลลิค ซึ่งอยู่ในสภาพสารละลายถูกทำให้สลายตัวได้โดยใช้ Acid Hydrolysis ที่อุณหภูมิสูงและได้ผลิตภัณฑ์คือกรดจิบเบอเรลลี-นิค (Gibberellenic Acid) และกรดจิบเบอริค (Gibberic Acid)")
53
การหาปริมาณจิบเบอเรลลิน
1. ใช้วิธีโครมาโตกราฟ เช่น GC หรือ Gas Chromatograph และ Paper Chromatograph 2. ใช้วิธี Bioassay โดยการที่จิบเบอเรลลินสามารถทำให้พืชแคระ (ข้าวโพดและถั่ว) เจริญเป็นต้นปกติได้ หรือโดยการที่จิบเบอเรลลินสามารถป้องกันการเกิดการเสื่อมสลาย (Senescence) หรือโดยหาปริมาณจิบเบอเรลลินจากการกระตุ้นให้เมล็ดข้าว-บาร์เลย์สร้างเอนไซม์ แอลฟา อะมัยเลส ( -amylase) ในอาหารสำรอง
 เจริญเป็นต้นปกติได้ หรือโดยการที่จิบเบอเรลลินสามารถป้องกันการเกิดการเสื่อมสลาย (Senescence) หรือโดยหาปริมาณจิบเบอเรลลินจากการกระตุ้นให้เมล็ดข้าว-บาร์เลย์สร้างเอนไซม์ แอลฟา อะมัยเลส ( -amylase) ในอาหารสำรอง.")
54
จิบเบอเรลลินสามารถเคลื่อนย้ายหรือเคลื่อนที่ในพืชได้ทั้งทางเบสิพีตัล และอะโครพีตัล และการเคลื่อนที่ไม่มีโพลาริตี้ การเคลื่อนย้ายเกิดขึ้นทั้งในส่วนของท่ออาหารและท่อน้ำ จิบเบอเรลลินจากยอดอ่อนลงมาสู่ส่วนล่างของลำต้นนั้นไม่ได้เกิดในท่อน้ำ ท่ออาหารเพราะส่วนของยอดอ่อนเป็นส่วนที่ดึงอาหารและธาตุอาหารให้เคลื่อนที่ขึ้นไปแบบอะโครพีตัล ดังนั้นจิบเบอเรลลิน จึงไม่ได้เคลื่อนที่ทางท่ออาหารและยังไม่ทราบวิถีการเคลื่อนที่แน่ชัด

55
กลไกในการทำงานของจิบเบอเรลลิน
ระดับของกิจกรรมของเอนไซม์หลายชนิดมีผลกระทบจากปริมาณของจิบเบอเรลลิน เอนไซม์ซึ่งมีกิจกรรมเพิ่มขึ้นเมื่อได้รับจิบเบอเรลลิน คือ เอนไซม์ แอลฟาและเบตา-อะมัยเลส ( และ -amylase) โปรตีเอส (Protease) และไรโบนิวคลีเอส (Ribonuclease) ซึ่งพบในเมล็ดข้าวบาร์เลย์ซึ่งกำลังงอก
 โปรตีเอส (Protease) และไรโบนิวคลีเอส (Ribonuclease) ซึ่งพบในเมล็ดข้าวบาร์เลย์ซึ่งกำลังงอก.")
56
ระดับกิจกรรมของเอนไซม์ซึ่งถูกควบคุมโดยจิบเบอ-เรลลิน ทำกันมากในเอนไซม์ แอลฟา อะมัยเลส ในเมล็ดข้าวบาร์เลย์ ในเมล็ดข้าวบาร์เลย์ที่แห้งที่ยังไม่ดูดซับน้ำจะไม่มีเอนไซม์แอลฟา อะมัยเลส ปรากฏอยู่เอนไซม์นี้จะปรากฏขึ้นและปลดปล่อยออกมาจากชั้นของอะลีโรนของเมล็ด เป็นการตอบสนองต่อจิบเบอ-เรลลินซึ่งสังเคราะห์จากต้นอ่อนที่กำลังงอก เนื้อเยื่อชั้นอะลีโรนซึ่งแยกจากเมล็ดที่ไม่งอกจะมีกิจกรรมของแอลฟา อะมัยเลส น้อยมาก

57
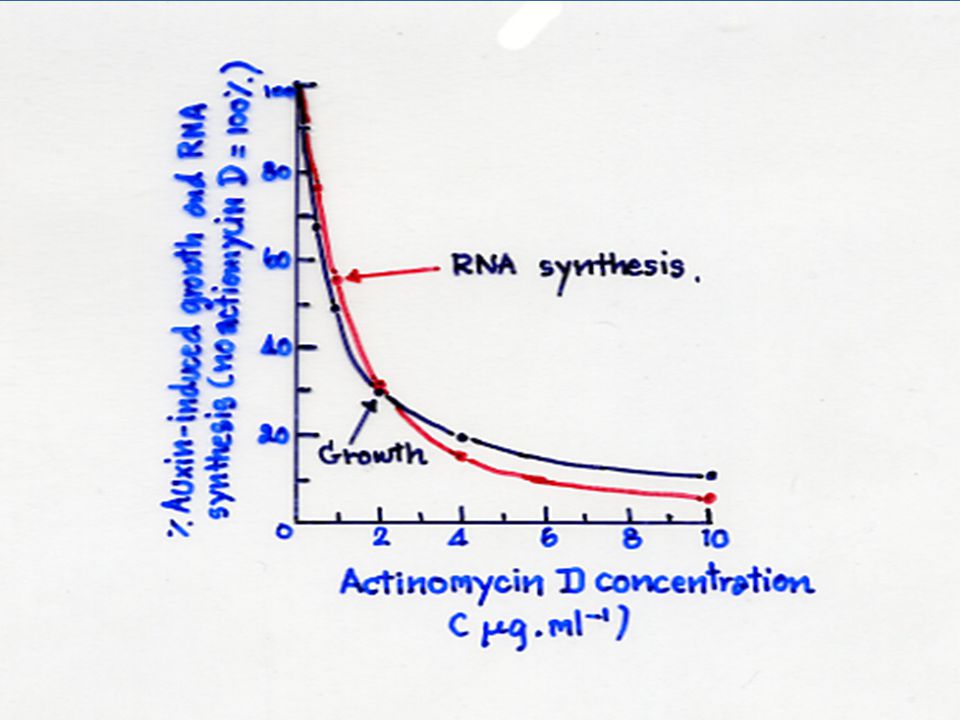
จิบเบอเรลลินควบคุมกิจกรรมของแอลฟา อะมัยเลส ผ่านทางการสังเคราะห์ RNA สารชะงักการสังเคราะห์ RNA เช่น แอคติโนมัยซิน-ดี (Actinomycin-D) จะชะงักกระบวนการกระตุ้นการสังเคราะห์ RNA 2-3 ชั่วโมง หลังจากเติมจิบเบอเรลลิน ในขณะที่สารชะงักการสังเคราะห์โปรตีน เช่น ไซโคลเฮคซิไมด์ (Cycloheximide) จะระงับการปรากฏของกิจกรรมของแอลฟา อะมัยเลส หลังจากช่วง "lag" เริ่มต้น
 จะชะงักกระบวนการกระตุ้นการสังเคราะห์ RNA 2-3 ชั่วโมง หลังจากเติมจิบเบอเรลลิน ในขณะที่สารชะงักการสังเคราะห์โปรตีน เช่น ไซโคลเฮคซิไมด์ (Cycloheximide) จะระงับการปรากฏของกิจกรรมของแอลฟา อะมัยเลส หลังจากช่วง lag เริ่มต้น")
58
บทบาทของจิบเบอเรลลินที่มีต่อพืช
1. กระตุ้นการเจริญเติบโตของพืชทั้งต้น 2. กระตุ้นการงอกของเมล็ดที่พักตัวและตาที่พักตัว 3. การแทงช่อดอก การออกดอกของพืชเกี่ยวข้องกับปัจจัยหลายอย่าง เช่น อายุ และสภาพแวดล้อม จิบเบอเรลลินสามารถแทนความต้องการวันยาวในพืชบางชนิดได้ และยังสามารถทดแทนความต้องการอุณหภูมิต่ำ (Vernalization) ในพืชพวกกะหล่ำปลี และแครอท
 ในพืชพวกกะหล่ำปลี และแครอท.")
59
4. จิบเบอเรลลิน สามารถกระตุ้นการเคลื่อนที่ของอาหารในเซลล์สะสมอาหารหลังจากที่เมล็ดงอกแล้ว
5. กระตุ้นให้เกิดผลแบบ Parthenocarpic ในพืชบางชนิด เปลี่ยนรูปร่างของใบพืชบางชนิด เช่น English Ivy และทำให้พืชพัฒนาการเพื่อทนความเย็นได้ 6. จิบเบอเรลลินมักเร่งให้เกิดดอกตัวผู้

61
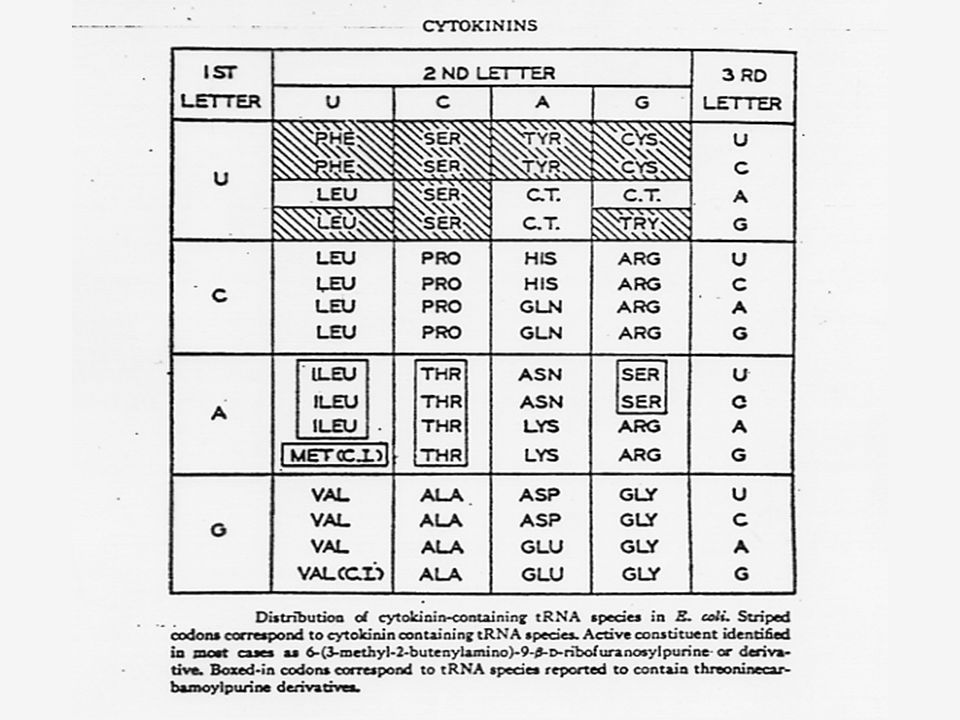
ไซโตไคนิน (Cytokinins)
1920 Haberlandt ได้แสดงให้เห็นว่ามีสารชนิดหนึ่งเกิดอยู่ในเนื้อเยื่อพืชและกระตุ้นให้เนื้อเยื่อพาเรนไคมาในหัวมันฝรั่งกลับกลายเป็นเนื้อเยื่อเจริญได้ ซึ่งแสดงว่าสารชนิดนี้สามารถกระตุ้นให้มีการแบ่งเซลล์ ต่อมามีการพบว่าน้ำมะพร้าวและเนื้อเยื่อของหัวแครอทมีคุณสมบัติในการกระตุ้นการแบ่งเซลล์เช่นกัน

62
ไซโตไคนิน (Cytokinins)
Skoog และ Steward ทำการทดลองในสหรัฐอเมริกา โดยศึกษาความต้องการสิ่งที่ใช้ในการเจริญเติบโตของกลุ่มก้อนของเซลล์ (Callus) ซึ่งเป็นเซลล์ที่แบ่งตัวอย่างรวดเร็ว แต่ไม่มีการเปลี่ยนแปลงทางคุณภาพเกิดขึ้นของ pith จากยาสูบและรากของแครอท
 ซึ่งเป็นเซลล์ที่แบ่งตัวอย่างรวดเร็ว แต่ไม่มีการเปลี่ยนแปลงทางคุณภาพเกิดขึ้นของ pith จากยาสูบและรากของแครอท.")
63
ไซโตไคนิน (Cytokinins)
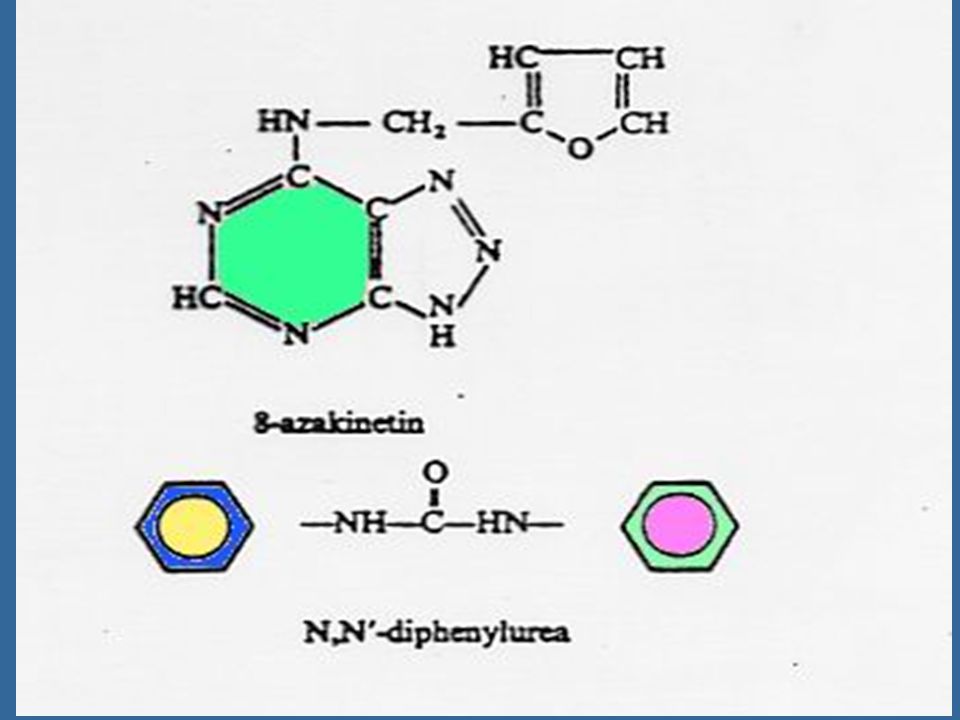
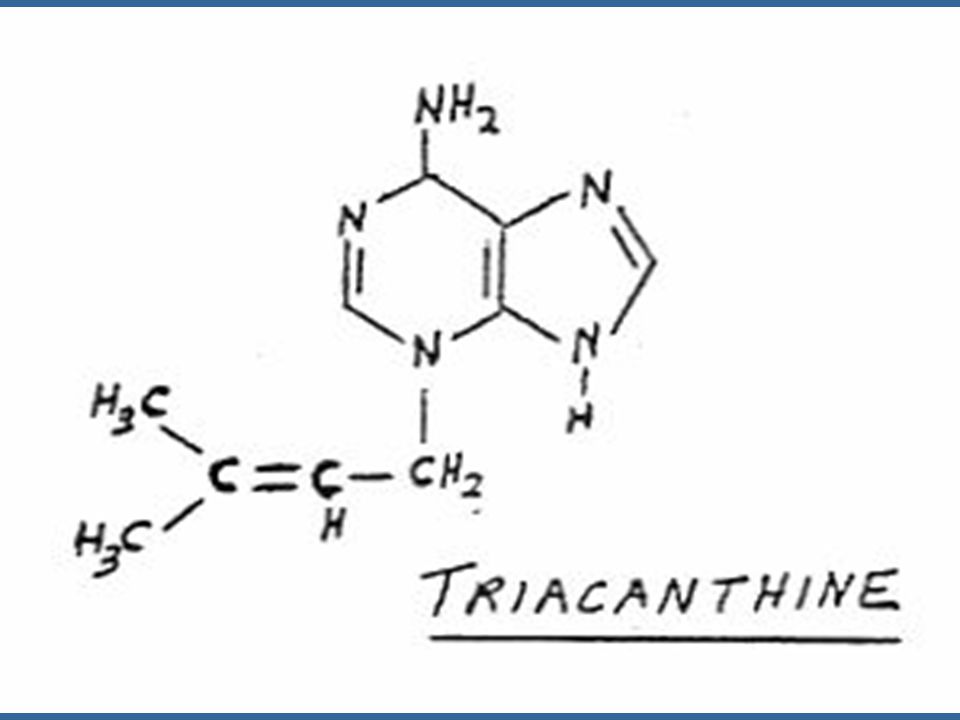
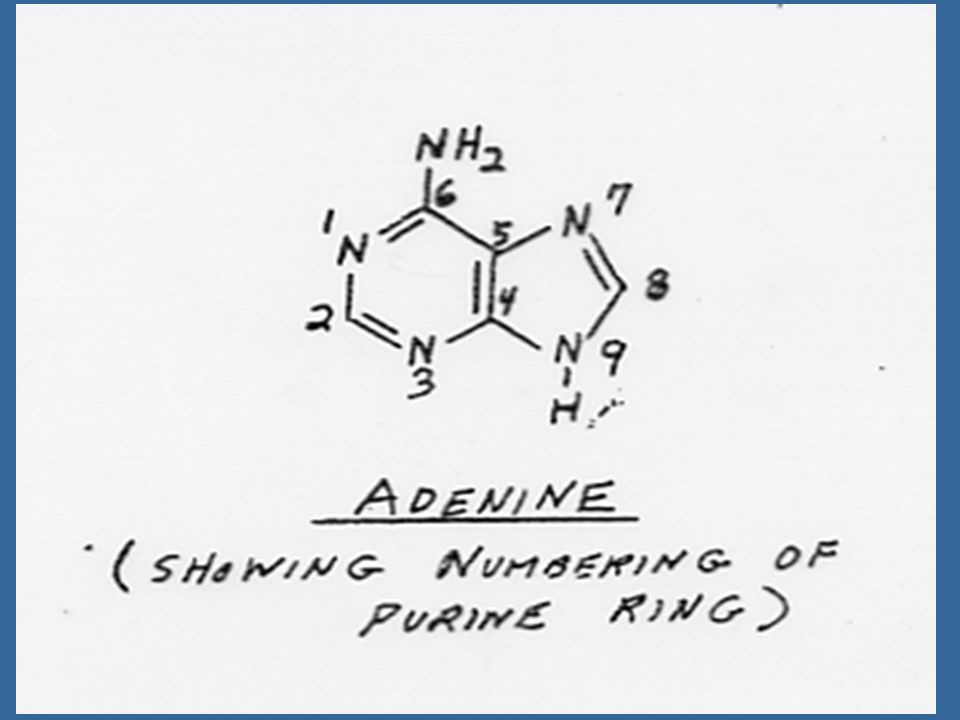
Miller ได้แยกสารอีกชนิดหนึ่งซึ่งมีคุณสมบัติคล้ายคลึงแต่มีประสิทธิภาพดีกว่าอะดีนีน ซึ่งได้จากการสลายตัวของ DNA ของสเปิร์มจากปลาแฮร์ริง สารชนิดนี้ คือ 6-(furfuryl-amino) purine ซึ่งมีสูตรโครงสร้างคล้ายอะดีนีน เนื่องจากสารชนิดนี้สามารถกระตุ้นให้เกิดการแบ่งเซลล์โดยร่วมกับ ออกซิน จึงได้รับชื่อว่าไคเนติน (Kinetin)
 purine ซึ่งมีสูตรโครงสร้างคล้ายอะดีนีน เนื่องจากสารชนิดนี้สามารถกระตุ้นให้เกิดการแบ่งเซลล์โดยร่วมกับ ออกซิน จึงได้รับชื่อว่าไคเนติน (Kinetin)")
64
สารสังเคราะห์ที่มีกิจกรรมของไซโตไคนินสูงที่สุดคือ เบนซิลอะดีนีน (Benzyladenine หรือ BA) และเตตระไฮโดรไพรานีลเบนซิลอะดีนีน (tetrahydropyranylbenzyladenine หรือ PBA)
 และเตตระไฮโดรไพรานีลเบนซิลอะดีนีน (tetrahydropyranylbenzyladenine หรือ PBA)")
65
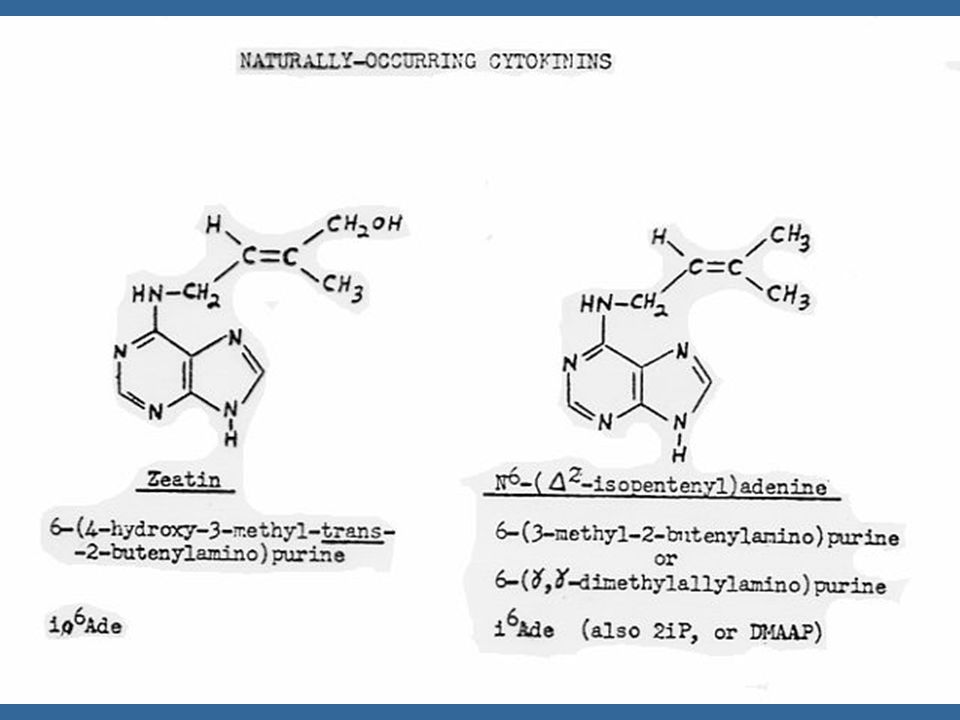
ไซโตไคนินที่เกิดในธรรมชาติเป็นสารประกอบพิวรีน ในปี Letham ได้แยกไซโตไคนินชนิดหนึ่งจากเมล็ดข้าวโพดหวาน และพบว่า เป็นสาร 6-(4-hydroxy-3-methyl but-2-enyl) aminopurine ซึ่ง Letham ได้ตั้งชื่อว่า ซีเอติน (Zeatin)
 aminopurine ซึ่ง Letham ได้ตั้งชื่อว่า ซีเอติน (Zeatin)")
66
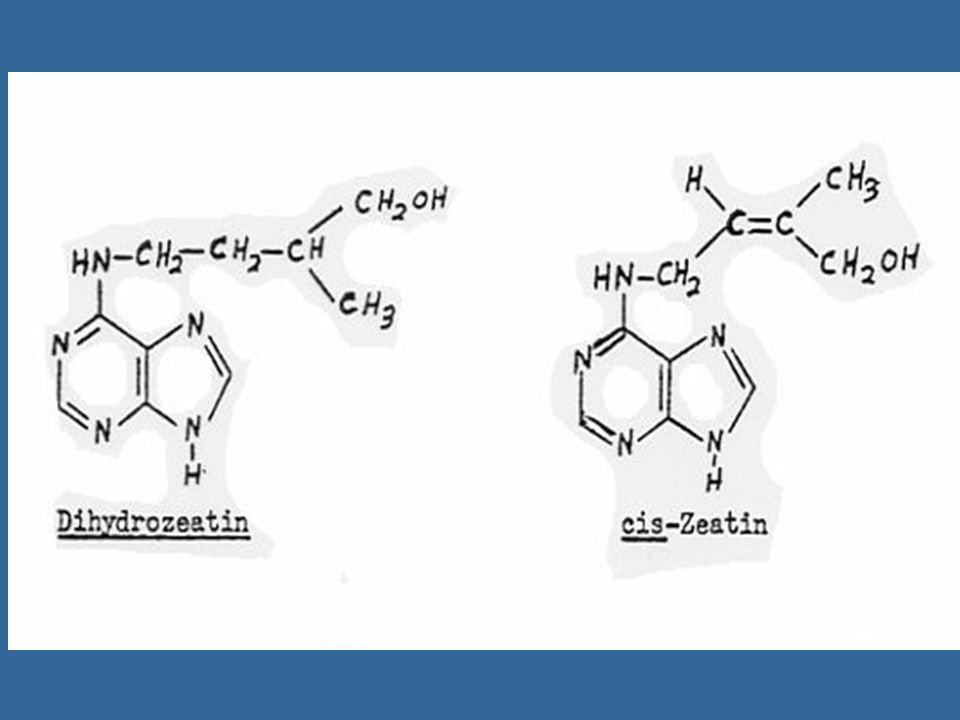
นับตั้งแต่มีการแยกไซโตไคนินชนิดแรกคือซีเอตินแล้ว ก็มีการค้นพบไซโตไคนิน อีกหลายชนิดซึ่งทุกชนิดเป็นอนุพันธ์ของอะดีนีน คือ เป็น 6-substituted amino purines ซีเอตินเป็นไซโต-ไคนินธรรมชาติซึ่งมีประสิทธิภาพสูงที่สุด

71
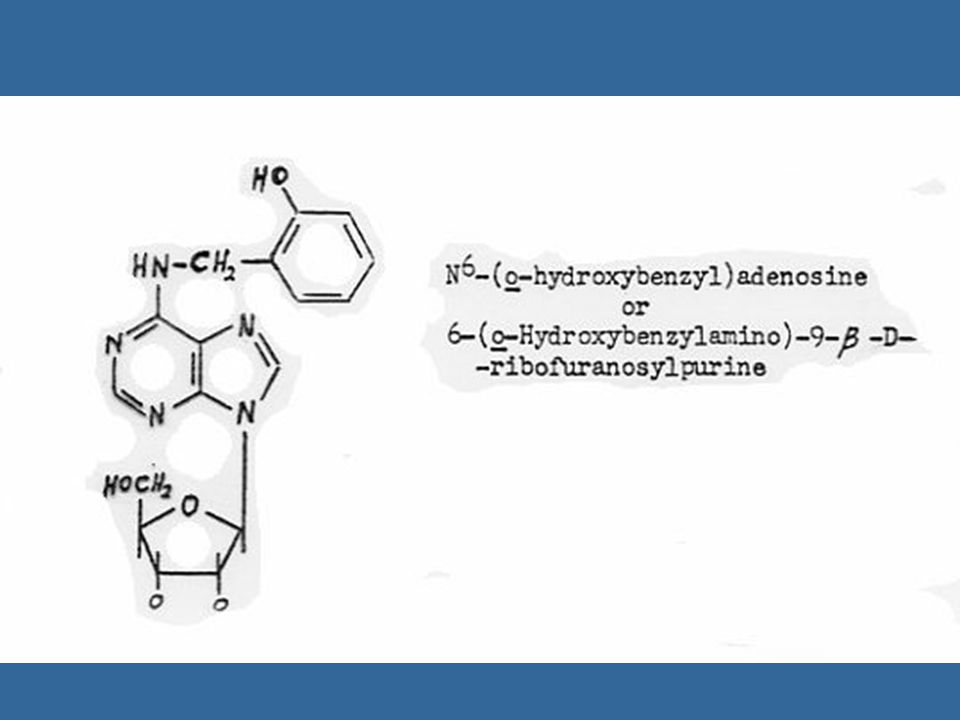
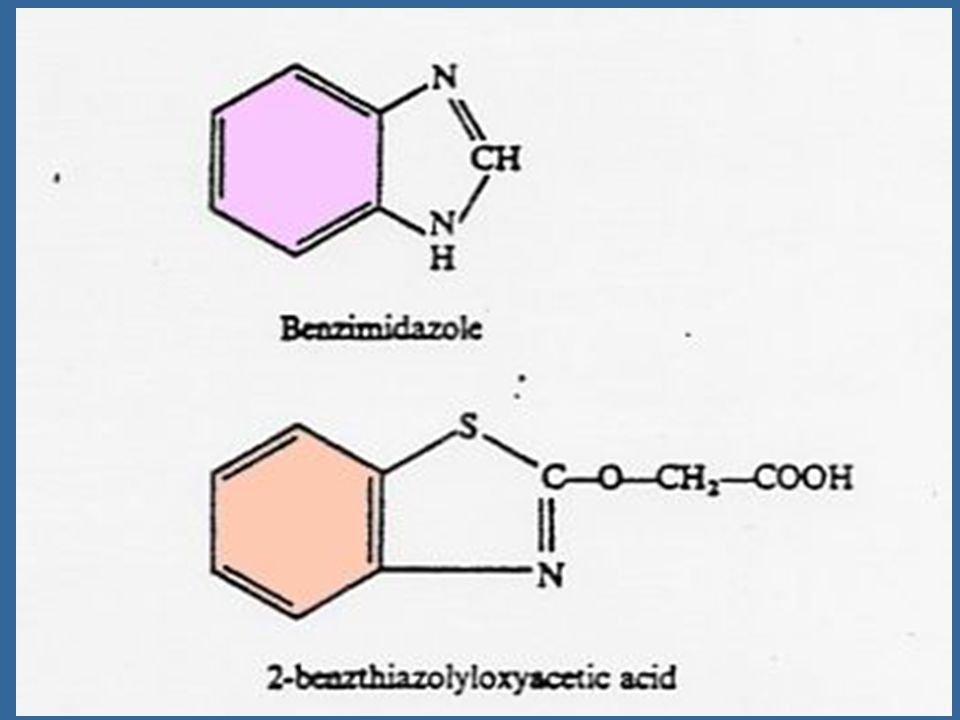
Non-purine Compounds with Cytokinin-like Activity

77
ไซโตไคนิน รองศาสตราจารย์ ดร. ดนัย บุณยเกียรติ
รองศาสตราจารย์ ดร. ดนัย บุณยเกียรติ ภาควิชาพืชสวน คณะเกษตรศาสตร์ มหาวิทยาลัยเชียงใหม่

78
การสังเคราะห์ไซโตไคนิน
การสังเคราะห์ไซโตไคนินต้นพืชเกิดโดยการ substitution ของ side chian บนคาร์บอนอะตอมที่6 ของอะดีนีน ซึ่ง side chian ของไซไคนีนในสภาพธรรมชาติประกอบด้วยคาร์บอน 5 อะตอม จึงเป็นการชี้ให้เห็นว่าเกิดมาจากวิถีการสังเคราะห์ไอโซพรีนอยด์ (Isopreoid) ต่อมาพบว่า กลุ่มของไซโตไคนินเกิดขึ้นบน t-RNA ได้ และเมื่อใช้เมวาโลเนต (Mavalonate หรือ MVA) ที่มีสารกัมมันตรังสีจะสามารถไปรวมกับกลุ่มอะดีนีนของ t-RNA เกิดเป็นไดเมทธิลอัลลิล (Dimethylallyl side chian) เกาะด้านข้าง ในเชื้อรา Rhizopus นั้น Dimethylallyl adenine สามารถเปลี่ยนไปเป็น Zeatin ได้ จึงคาดว่า Zeatin อาจจะเกิดจากออกซิ-ไดซ์ Dimethylallal adenine
 ต่อมาพบว่า กลุ่มของไซโตไคนินเกิดขึ้นบน t-RNA ได้ และเมื่อใช้เมวาโลเนต (Mavalonate หรือ MVA) ที่มีสารกัมมันตรังสีจะสามารถไปรวมกับกลุ่มอะดีนีนของ t-RNA เกิดเป็นไดเมทธิลอัลลิล (Dimethylallyl side chian) เกาะด้านข้าง ในเชื้อรา Rhizopus นั้น Dimethylallyl adenine สามารถเปลี่ยนไปเป็น Zeatin ได้ จึงคาดว่า Zeatin อาจจะเกิดจากออกซิ-ไดซ์ Dimethylallal adenine.")
79
พบไซโตไคนินมากในผลอ่อนและเมล็ด ในใบอ่อนและปลายรากซึ่งไซโตไคนินอาจจะสังเคราะห์ที่บริเวณดังกล่าวหรืออาจจะเคลื่อนย้ายมาจากส่วนอื่นๆ ในรากนั้นมีหลักฐานที่ชี้ชัดให้เห็นว่าไซโตไคนินสังเคราะห์ที่บริเวณนี้ได้เพราะเมื่อมีการตัดรากหรือลำต้น พบว่าของเหลวที่ไหลออกมาจากท่อน้ำจะปรากฏไซโตไคนินจากส่วนล่างขึ้นมา ติดต่อกันถึง 4 วัน ซึ่งอาจจะเป็นไปได้ว่าไซโตไคนินสังเคราะห์จากที่รากแล้วส่งไปยังส่วนอื่นๆ โดยทางท่อน้ำ หลักฐานที่แสดงว่าสังเคราะห์ที่ส่วนอื่นยังไม่พบและการเคลื่อนย้ายของไซโต-ไคนินจากส่วนอ่อน เช่น ใบ เมล็ด ผล ยังเกิดไม่ดีและไม่มาก

80
การสลายตัวของไซโตไคนิน
ไซโตไคนินสามารถถูกทำลายโดยการออกซิเดชั่น ทำให้ side chian อะดีนีน ติดตามด้วยการทำงานของเอ็นไซม์ แซนทีออกซิเดส (Xanthine Oxidase) ซึ่งสามารถออกซิไดซ์ พิวรีนเกิดเป็นกรดยูริค (Uric Acid) และกลายเป็นยูเรียไปในที่สุด อย่างไรก็ตามในใบพืชไซโตไคนินอาจจะถูกเปลี่ยนไปเป็นกลูโคไซด์ โดยน้ำตาลกลูโคสจะไปเกาะกับตำแหน่งที่ 7 ของอะดีนีนเกิดเป็น 7-กลูโคซีลไซโตไคนิน (7-glucosylcyto- kinins) หน้าที่ของไซโตไคนิน กลูโคไซด์ยังไม่ทราบแน่ชัดนัก อาจจะเป็น “detoxification” ซึ่งไม่เกี่ยวข้องกับกิจกรรมทางเมตาบอลิสม์หรืออาจจะเป็นรูปที่ไซโตไคนินอาจจะถูกปลดปล่อยออกมาในบางสภาวะได้ จากการศึกษาโดยใช้ Radioactive BA พบว่าสามารถสลายตัวกลายเป็นกรดยูริคแล้วอาจจะรวมกับ RNA ได้
 ซึ่งสามารถออกซิไดซ์ พิวรีนเกิดเป็นกรดยูริค (Uric Acid) และกลายเป็นยูเรียไปในที่สุด อย่างไรก็ตามในใบพืชไซโตไคนินอาจจะถูกเปลี่ยนไปเป็นกลูโคไซด์ โดยน้ำตาลกลูโคสจะไปเกาะกับตำแหน่งที่ 7 ของอะดีนีนเกิดเป็น 7-กลูโคซีลไซโตไคนิน (7-glucosylcyto- kinins) หน้าที่ของไซโตไคนิน กลูโคไซด์ยังไม่ทราบแน่ชัดนัก อาจจะเป็น detoxification ซึ่งไม่เกี่ยวข้องกับกิจกรรมทางเมตาบอลิสม์หรืออาจจะเป็นรูปที่ไซโตไคนินอาจจะถูกปลดปล่อยออกมาในบางสภาวะได้ จากการศึกษาโดยใช้ Radioactive BA พบว่าสามารถสลายตัวกลายเป็นกรดยูริคแล้วอาจจะรวมกับ RNA ได้")
84
การเคลื่อนที่ของไซโตไคนิน
ยังไม่มีหลักฐานว่าเคลื่อนที่อย่างไรแน่ จากการทดลองทราบว่าระบบรากเป็นส่วนสำคัญในการส่งไซโตไคนินไปยังใบ และป้องกันการเสื่อมสลายของใบก่อนระยะอันสมควรเป็นหลักฐานที่สำคัญที่ชี้ให้เห็นว่า ไซโตไคนินมีการเคลื่อนที่ขึ้นสู่ยอด ในทางตรงกันข้ามไซโตไคนินซึ่งพบที่ผลซึ่งกำลังเจริญเติบโตไม่เคลื่อนที่ไปส่วนอื่นเลย ในทำนองเดียวกันจากการศึกษากับการให้ไซโตไคนินจากภายนอก เช่น ให้ไคเนติน พบว่าจะไม่เคลื่อนย้ายเป็นเวลานาน

85
ในการให้ไซโตไคนินกับตาข้างเพื่อกำจัด Apical dominance นั้น พบว่าไซโตไคนินจะไม่เคลื่อนที่เลยเป็นระยะเวลานานมาก ในการทดลองกับ BA พบว่า BA สามารถเคลื่อนที่ผ่านก้านใบและลักษณะแบบ Polar เหมือนกับออกซิเจนในทุการศึกษาพบว่า ไซโตไคนินในใบจะไม่เคลื่อนที่รวมทั้งในผลอ่อนด้วย ส่วนผลของรากในการควบคุมการเจริญเติบโตของส่วนเหนือดินอาจจะอธิบายได้ถึงไซโตไคนินที่เคลื่อนที่ในท่อน้ำ ซึ่งพบเสมอในการทดลองว่าไซโตไคนินสามารถเคลื่อนที่จากส่วนรากไปสู่ยอด แต่การเคลื่อนที่แบบ Polar ยังไม่เป็นที่ยืนยันการเคลื่อนที่ของไซโตไคนินในพืชยังมีความขัดแย้งกันอยู่บ้าง

86
การหาปริมาณของไซโตไคนิน
1. ใช้ Toobacco callus test 2. Leaf senescence test

87
ไซโตไคนินที่เกิดในธรรมชาติทั้งหมดเป็นสารที่เป็นอะดีนีน มี side chain ซึ่งมีคาร์บอน 5 อะตอมเกาะอยู่ที่ตำแหน่งที่ 6 ของแหวนอะดีนีน ซึ่งจากการศึกษาพบว่า เมื่อ side chain ไม่เป็นวงแหวนจำนวนคาร์บอนอะตอมที่เหมาะสมที่สุดคือ 5 อะตอม การเพิ่มหรือการลดขนาดของ side chain จะลดกิจกรรมทางสรีรวิทยาของไซโตไคนินลง แต่ไม่ทำให้กิจกรรมหมดไปการปรากฏของแขนแบบ double bond ขึ้นที่ side chain จะเพิ่มประสิทธิภาพของไซโตไคนินให้มากขึ้น

88
กลไกการทำงานของไซโตไคนิน
ไซโตไคนินมีบทบาทสำคัญคือควบคุมการแบ่งเซลล์ และไซโตไคนินที่เกิดในสภาพธรรมชาตินั้นเป็นอนุพันธุ์ของอะดีนีนทั้งสิ้น ดังนั้นงานวิจัยเกี่ยวกับกลไกการทำงานจึงมีแนวโน้มในความสัมพันธ์กับกรดนิวคลีอิค กลไกการทำงานของไซโตไคนินยังไม่เด่นชัดเหมือนกับออกซิน และจิบเบอเรลลิน แต่ไซโตไคนินมีผลให้เกิดการสังเคราะห์ RNA และโปรตีนมากขึ้นในเซลล์พืช ผลการทดลองรายงานกล่าวว่า หลังจากให้ไซโตไคนินกับเซลล์พืชแล้วจะเพิ่มปริมาณของ m-RNA , t-RNA และ r-RNA

89
ไซโตไคนินอาจจะทำงานโดยควบคุมกิจกรรมของเอ็นไซม์โดยตรงมากกว่าที่เกี่ยวกับการสังเคราะห์เอนไซม์ ไซโตไคนินมีอิทธิพลต่อเอนไซม์หลายชนิด เช่น ไคเนส (Kinases) ที่ใช้ในกระบวนการหายใจ นอกจากนั้นกิจกรรมของเอนไซม์ที่ใช้ในกระบวนการสังเคราะห์แสงก็เพิ่มขึ้น
 ที่ใช้ในกระบวนการหายใจ นอกจากนั้นกิจกรรมของเอนไซม์ที่ใช้ในกระบวนการสังเคราะห์แสงก็เพิ่มขึ้น")
90
ผลของไซโตไคนิน 1. กระตุ้นให้เกิดการแบ่งเซลล์และการเปลี่ยนแปลงทางคุณภาพใน tissue culture โดยต้องใช้ร่วมกับ Auxin ในการเลี้ยงเนื้อเยื่อพืชนั้นหากฮอร์โมน ไซโตไคนินมากกว่าออกซินจะทำให้เนื้อเยื่อนั้นเจริญเป็น ตา ใบ และลำต้น แต่ถ้าหากสัดส่วนของออกซินมากขึ้นกว่าไซโตไคนินจะทำให้เนื้อเยื่อนั้นสร้างรากขึ้นมา การ differentiate ของตาในการเพาะเลี้ยงเนื้อเยื่อจาก Callus จากส่วนของลำต้นนั้น auxin จะระงับ และไซโตไคนินนั้นจะกระตุ้นการเกิดและต้องมีความสมดุลระหว่างไซโตไคนินและออกซินชิ้นเนื้อเยื่อจึงจะสร้างตาได้

91
ผลของไซโตไคนิน 2. ชะลอกระบวนการเสื่อมสลาย เช่น กรณีของใบที่เจริญเต็มที่แล้วถูกตัดออกจากต้น คลอโรฟิลล์ RNA และโปรตีนจะเริ่มสลายตัวเร็วกว่าใบที่ติดอยู่กับต้น 3. ทำให้ตาข้างแตกออกมาหรือกำจัดลักษณะ Apical Dominance ได้ การเพิ่มไซโตไคนินให้กับตาข้างจะทำให้แตกออกมาเป็นใบได้ ทั้งนี้เพราะตาข้างจะดึงอาหารมาจากส่วนอื่นทำให้ตาข้างเจริญได้

92
ผลของไซโตไคนิน 4. ทำให้ใบเลี้ยงคลี่ขยายตัว
5.ทำให้เกิดการสร้างคลอโรพลาสต์มากขึ้น ซึ่งเป็นการเปลี่ยนแปลงทางคุณภาพอย่างหนึ่ง เช่น เมื่อ Callus ได้รับแสงและไซโตไคนิน Callus จะกลายเป็นสีเขียว 6. ทำให้พืชทั้งต้นเจริญเติบโต 7.กระตุ้นการงอกของเมล็ดพืชบางชนิด

93
กรดแอบซิสิค (Abscisic Acid) หรือ ABA
 หรือ ABA")
94
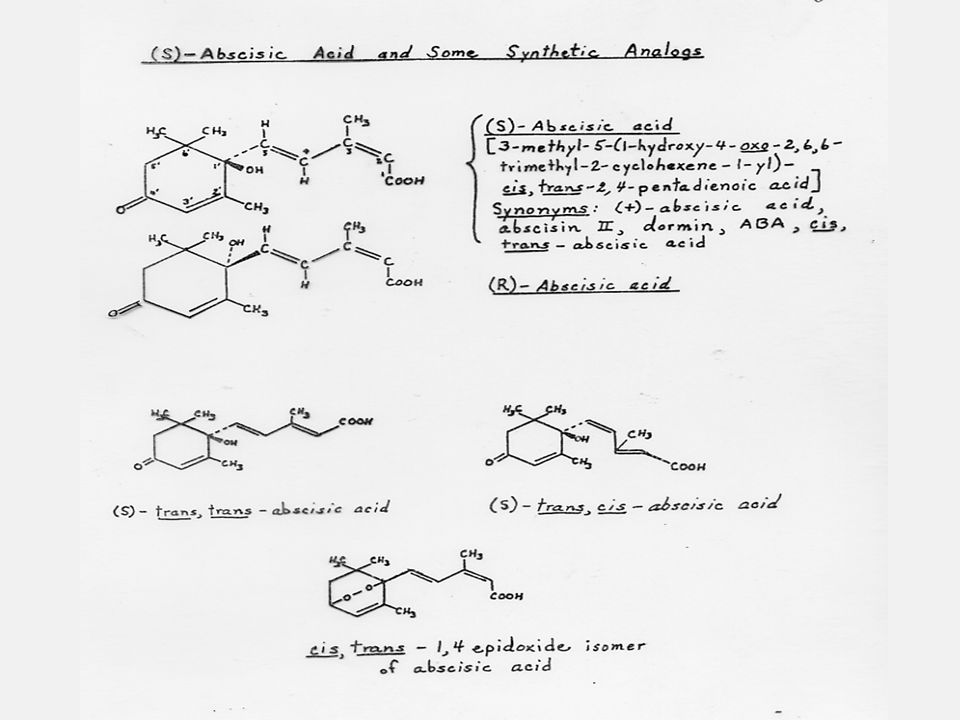
กรดแอบซิสิค (Abscisic Acid) หรือ ABA
โมเลกุลของ ABA ประกอบด้วย asymmetric carbon atom จึงสามารถแสดงลักษณะของ optical isomerism ได้ อย่างไรก็ตามในสภาพธรรมชาติ ABA จะเกิดเพียงชนิด (+) enantiomorph เท่านั้น ABA ยังแสดงลักษณะ geometric isomerism ได้ด้วย side chain จะเป็น trans รอบ ๆคาร์บอนตำแหน่งที่ 5 เสมอ แต่โมเลกุลสามารถเป็นได้ทั้ง cis- หรือ trans รอบๆคาร์บอนตำแหน่งที่2 ABA ส่วนใหญ่ที่พบในพืชจะเป็น (+)-2-cis ABA แม้ว่าจะพบ (+)-2-trans ABA บ้างแต่น้อยมาก ดังนั้นรูป (+)-2-cis ของ ABA จึงมักหมายถึง ABA ทั่วไป ABA ถูกแยกออกจากพืชหลายชนิดทั้งแองจิโอสเปิร์มส์ จิมโน สเปิร์มส์ เฟินและมอส (Angiosperms,Gymnosperms,Ferns และ Mosses
 enantiomorph เท่านั้น ABA ยังแสดงลักษณะ geometric isomerism ได้ด้วย side chain จะเป็น trans รอบ ๆคาร์บอนตำแหน่งที่ 5 เสมอ แต่โมเลกุลสามารถเป็นได้ทั้ง cis- หรือ trans รอบๆคาร์บอนตำแหน่งที่2 ABA ส่วนใหญ่ที่พบในพืชจะเป็น (+)-2-cis ABA แม้ว่าจะพบ (+)-2-trans ABA บ้างแต่น้อยมาก ดังนั้นรูป (+)-2-cis ของ ABA จึงมักหมายถึง ABA ทั่วไป. ABA ถูกแยกออกจากพืชหลายชนิดทั้งแองจิโอสเปิร์มส์ จิมโน สเปิร์มส์ เฟินและมอส (Angiosperms,Gymnosperms,Ferns และ Mosses.")
96
การสังเคราะห์ ABA ABA เป็นสารประกอบที่มีคาร์บอน 15 อะตอมและเป็นเซสควิเทอร์พีนอยด์ ดังนั้นวิถีการสังเคราะห์จึงใช้วิถีของการสังเคราะห์สารเทอร์พีนอยด์ โดยสังเคราะห์จากกรดเมวาโลนิคเปลี่ยนไปจนเป็นฟาร์นีซีล ไพโรฟอสเฟต (Farnaesyl pyrophosphate) ABA สังเคราะห์ในใบแก่และผลแก่ และจะถูกกระตุ้นให้สังเคราะห์เมื่อขาดน้ำ โดยการสังเคราะห์เกิดในคลอโรพลาสต์ ดังนั้น ใบ ลำต้น และผลไม้สีเขียวจึงสังเคราะห์ ABA ได้ นอกจากนั้นแหล่งอาหารสำรองและรากของข้าวสาลีก็สามารถสังเคราะห์ ABA ได้ เมื่อเกิดการสังเคราะห์ ABAแล้ว ABA จะเคลื่อนย้ายจากใบไปสู่ส่วนอื่นๆ เช่น ยอด และไประงับการเจริญที่บริเวณนั้น อาจจะกระตุ้นให้เกิดการพักตัวของตาในเมล็ดอาจจะมีการสังเคราะห์ ABA ได้บ้าง หรือ อาจจะเป็น ABA ซึ่งส่งจากใบ แต่ในเมล็ดมักจะมีปริมาณของ ABA อยู่มาก
 ABA สังเคราะห์ในใบแก่และผลแก่ และจะถูกกระตุ้นให้สังเคราะห์เมื่อขาดน้ำ โดยการสังเคราะห์เกิดในคลอโรพลาสต์ ดังนั้น ใบ ลำต้น และผลไม้สีเขียวจึงสังเคราะห์ ABA ได้ นอกจากนั้นแหล่งอาหารสำรองและรากของข้าวสาลีก็สามารถสังเคราะห์ ABA ได้ เมื่อเกิดการสังเคราะห์ ABAแล้ว ABA จะเคลื่อนย้ายจากใบไปสู่ส่วนอื่นๆ เช่น ยอด และไประงับการเจริญที่บริเวณนั้น อาจจะกระตุ้นให้เกิดการพักตัวของตาในเมล็ดอาจจะมีการสังเคราะห์ ABA ได้บ้าง หรือ อาจจะเป็น ABA ซึ่งส่งจากใบ แต่ในเมล็ดมักจะมีปริมาณของ ABA อยู่มาก.")
97
การสังเคราะห์ ABA ระดับ ABA ภายในต้นพืชมีปริมาณขึ้นๆลงๆ ตามอัตราการเจริญพลังงานที่ทำงานได้ของน้ำในต้นพืช ในต้นพืชต้องมีการทำลาย ABA ด้วย แต่กลไกการทำลายยังไม่ทราบแน่ชัดนัก แต่พบว่าพบว่าถ้าให้ 14 C-ABA แก่พืช ABA ดังกล่าวจะเปลี่ยนไปเป็นกลูโคสเอสเตอร์ของ ABA อย่างรวดเร็ว (Glucocose Ester) ซึ่งค่อนข้างจะเสถียรในต้นพืชและยังมีกิจกรรมคล้ายคลึงกับ ABA ส่วนการสลายตัวของ ABA นั้น เกิดขึ้นโดยเกิดไฮดรอกซีเลชั่นและออกซินเดชั่นของกลุ่มเมทธิลที่เกาะอยู่กับวงแหวน
 ซึ่งค่อนข้างจะเสถียรในต้นพืชและยังมีกิจกรรมคล้ายคลึงกับ ABA ส่วนการสลายตัวของ ABA นั้น เกิดขึ้นโดยเกิดไฮดรอกซีเลชั่นและออกซินเดชั่นของกลุ่มเมทธิลที่เกาะอยู่กับวงแหวน.")
98
การหาปริมาณ ABA 1. Bioassay 2. ใช้ Gas chromatograph

99
ความสัมพันธ์ของโครงสร้างและกิจกรรมของ ABA
ทั้ง ABA ที่เกิดตามธรรมชาติ (+) type dextorotary และสังเคราะห์ (-) เป็น optical enantiomers กัน มีประสิทธิภาพต่อพืชเท่ากันแต่ 2-cis geometric isomer เท่านั้นที่มีคุณสมบัติของฮอร์โมน
 type dextorotary และสังเคราะห์ (-) เป็น optical enantiomers กัน มีประสิทธิภาพต่อพืชเท่ากันแต่ 2-cis geometric isomer เท่านั้นที่มีคุณสมบัติของฮอร์โมน.")
102
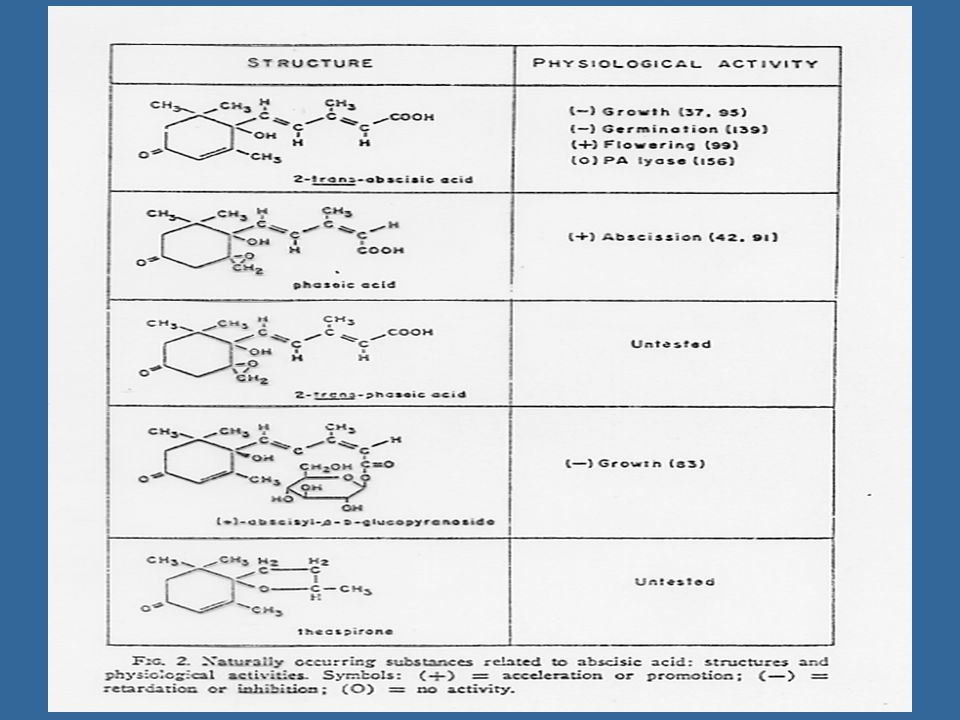
การสลายตัวของ ABA 1.เมื่อ ABA เปลี่ยนจากรูปของ cis ไปเป็น trans isomer ก็จะหมดคุณสมบัติในการทำให้พืชตอบสนองทางสรีรวิทยา 2. ถูกออกซิไดซ์กลายเป็น Phaseic acid 3. เกิดเป็นรูปที่รวมกับสารอื่น เช่น เกิด Glucose ester 4. เกิดเป็นสารประกอบกลุ่ม Methyl ester ซึ่งอาจจะยังมีผลต่อการตอบสนองของพืชบ้าง แต่น้อยลงและไม่กระตุ้นให้เกิดการปิดของปากใบ

103
กลไกในการทำงานของ ABA
เกี่ยวข้องกับการควบคุมเมตาบอลิสม์ของกรดนิวคลีอิค และการสังเคราะห์โปรตีน การปิดปากใบ ซึ่งเกิดภายในไม่กี่นาทีที่ได้รับ ABA นอกจากนั้นการยืดตัวของโคลีออพไทล์ที่ได้รับออกซินจะหยุดชะงักภายใน 2-3 นาที

104
การเคลื่อนที่ของ ABA เคลื่อนที่ทุกทิศทางโดยไม่มีโพลารีตี้ แต่ราก ABA อาจจะเคลื่อนที่ในลักษณะเบสิพีตัล

105
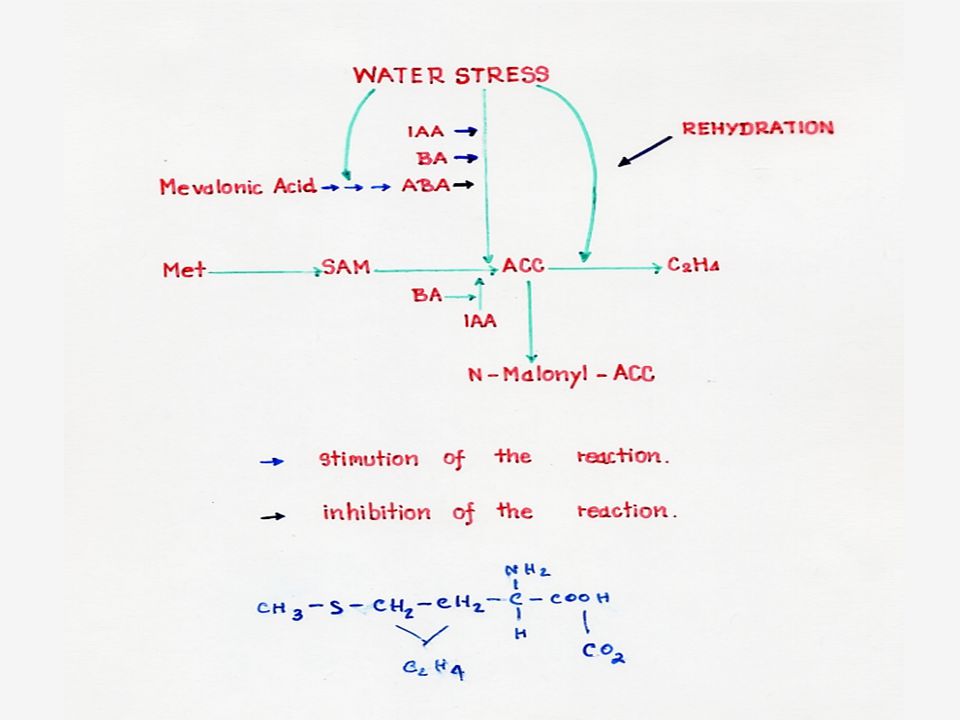
ความสัมพันธ์ของ ABA กับการขาดน้ำ
การขาดน้ำทำให้เยื่อหุ้มคลอโรพลาสต์มี Permeability เพิ่มขึ้น ส่งผลให้เกิดการรั่วไหลของ ABA จากคลอโรพลาสต์ซึ่งเป็นบริเวณที่เหมาะสม ABA ของใบปกติ การลดปริมาณของ ABA ในคลอโรพลาสต์จะกระตุ้นให้มีการสังเคราะห์ ABA เพิ่มขึ้น เมื่อพืชได้รับน้ำเยื่อหุ้มคลอโรพลาสต์จะไม่ยอมให้ ABA รั่วไหลออกมา ทำให้การหยุดสังเคราะห์ ABA ในคลอโรพลาสต์ ซึ่งเป็น Feedback mechanism

106
ABA กระตุ้นให้เกิดการปิดของปากใบได้เพราะ ABA จะระงับการผ่านของ K+ เข้าสู่ Guard cell และกระตุ้นให้ Matate รั่วไหลออกจาก Guard cell และนอกจากนั้น ABA ยังระงับการแลกเปลี่ยนประจุ H + และ K + ของ Guard cell ทำให้ปริมาณของ K + และ Malate ของ Guard cell มีน้อย ทำให้ค่า Water potential สูงขึ้น น้ำจึงไหลออกจาก Guard cell ทำให้ปากปิดได้

107
ผลของ ABA ต่อพืช 1. ลดการคายน้ำโดยกระตุ้นให้ปากใบปิด
2. กระตุ้นให้เกิดการพักตัวของตา 3. การร่วงของใบและดอก เช่น ในฝ้าย ผลแก่ที่ร่วงเองจะมี ABA สูงมาก 4. เร่งกระบวนการเสื่อมสภาพของใบ

108
เอทธิลีน (Ethylene) เอทธิลีนเป็นฮอร์โมนที่มีสภาพเป็นก๊าซซึ่งรู้จักกันมานานแล้ว จากการบ่มผลไม้ ในปี 1934 ได้มีการพิสูจน์ให้เห็นว่าเอทธิลีนเป็นก๊าซที่สังเคราะห์ขึ้นโดยพืช และสามารถเร่งกระบวนการสุกได้

109
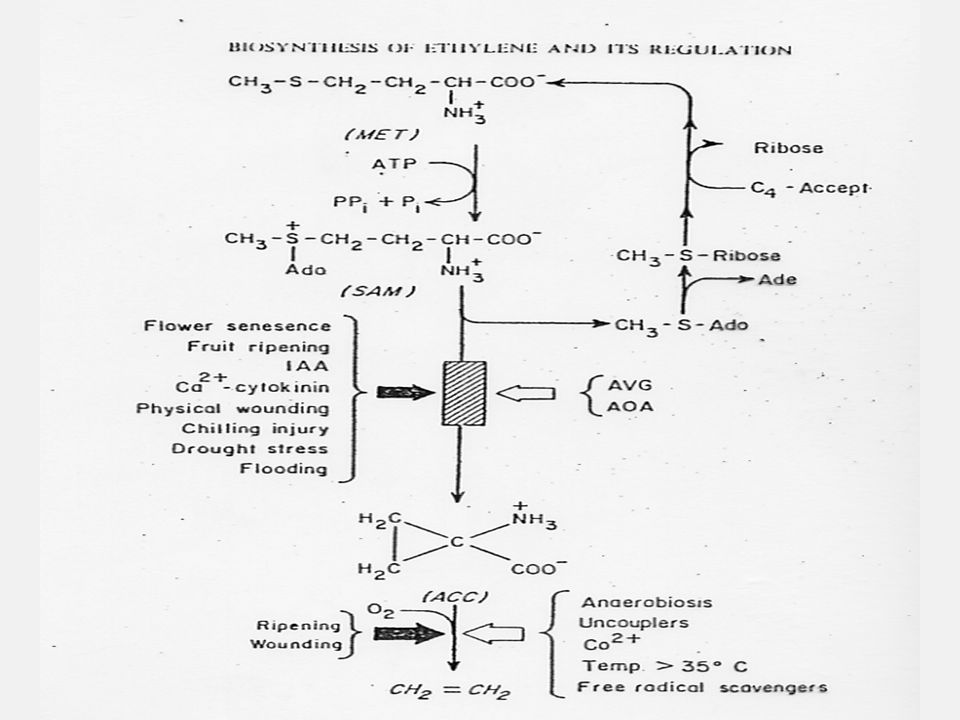
การสังเคราะห์เอทธิลีน
การสังเคราะห์เอทธิลีนเกิดโดยที่เมทไธโอนีนจะเปลี่ยนไปเป็น SAM (S-Adenosylmethionine) แล้วเปลี่ยนต่อไปเป็น ACC (1-aminocyclopropane-1-carboxylic acid) แล้วจึงเปลี่ยนเป็นเอทธิลีน โดยที่คาร์บอนอะตอมที่ 3 และ 4 ของเมทไธโอนีนจะกลายเป็นคาร์บอนของเอทธิลีน ในการสังเคราะห์นี้จะให้ก๊าซคาร์บอนไดออกไซด์ออกมาด้วย การสังเคราะห์ เอทธิลีนจะหยุดชะงักเมื่อบรรยากาศขาด O2 นอกจากนั้นยังมีสารระงับการสังเคราะห์เอทธิลีนชนิดอื่นๆ อีก เช่น AVG (Aminoethoxy vinyl glycine)
 แล้วเปลี่ยนต่อไปเป็น ACC (1-aminocyclopropane-1-carboxylic acid) แล้วจึงเปลี่ยนเป็นเอทธิลีน โดยที่คาร์บอนอะตอมที่ 3 และ 4 ของเมทไธโอนีนจะกลายเป็นคาร์บอนของเอทธิลีน ในการสังเคราะห์นี้จะให้ก๊าซคาร์บอนไดออกไซด์ออกมาด้วย การสังเคราะห์ เอทธิลีนจะหยุดชะงักเมื่อบรรยากาศขาด O2 นอกจากนั้นยังมีสารระงับการสังเคราะห์เอทธิลีนชนิดอื่นๆ อีก เช่น AVG (Aminoethoxy vinyl glycine)")
110
การสังเคราะห์เอทธิลีน
การสังเคราะห์เอทธิลีนเกิดโดยที่เมทไธโอนีนจะเปลี่ยนไปเป็น SAM (S-Adenosylmethionine) แล้วเปลี่ยนต่อไปเป็น ACC (1-aminocyclopropane-1-carboxylic acid) แล้วจึงเปลี่ยนเป็นเอทธิลีน โดยที่คาร์บอนอะตอมที่ 3 และ 4 ของเมทไธโอนีนจะกลายเป็นคาร์บอนของเอทธิลีน ในการสังเคราะห์นี้จะให้ก๊าซคาร์บอนไดออกไซด์ออกมาด้วย การสังเคราะห์ เอทธิลีนจะหยุดชะงักเมื่อบรรยากาศขาด O2 นอกจากนั้นยังมีสารระงับการสังเคราะห์เอทธิลีนชนิดอื่นๆ อีก เช่น AVG (Aminoethoxy vinyl glycine) เอทธิลีนสามารถถูกทำลายให้เป็นเอทธิลีนออกไซด์ (Ethylene oxide) ใน Vicia faba แต่กลไกในการทำลายนั้นยังไม่ทราบแน่ชัดนัก
 แล้วเปลี่ยนต่อไปเป็น ACC (1-aminocyclopropane-1-carboxylic acid) แล้วจึงเปลี่ยนเป็นเอทธิลีน โดยที่คาร์บอนอะตอมที่ 3 และ 4 ของเมทไธโอนีนจะกลายเป็นคาร์บอนของเอทธิลีน ในการสังเคราะห์นี้จะให้ก๊าซคาร์บอนไดออกไซด์ออกมาด้วย การสังเคราะห์ เอทธิลีนจะหยุดชะงักเมื่อบรรยากาศขาด O2 นอกจากนั้นยังมีสารระงับการสังเคราะห์เอทธิลีนชนิดอื่นๆ อีก เช่น AVG (Aminoethoxy vinyl glycine) เอทธิลีนสามารถถูกทำลายให้เป็นเอทธิลีนออกไซด์ (Ethylene oxide) ใน Vicia faba แต่กลไกในการทำลายนั้นยังไม่ทราบแน่ชัดนัก.")
111
การหาปริมาณของเอทธิลีน
การใช้คุณสมบัติทางฟิสิกส์ของเอทธิลีน ซึ่งอาจจะวัดโดย Gas Chromatograph

113
การเคลื่อนที่ของเอทธิลีน
เอทธิลีนจะไม่เคลื่อนที่ในส่วนต่าง ๆ ของพืชในปริมาณที่มากพอที่จะก่อให้เกิดการเปลี่ยนแปลงทางสรีรวิทยาได้ ทั้ง ๆ ที่เอทธิลีนไม่มีการเคลื่อนที่ในพืช แต่พบว่าระดับของเอทธิลีนในส่วนหนึ่งของพืชจะส่งผลให้เกิดการสังเคราะห์เอทธิลีนในส่วนอื่น ๆ ด้วย เช่น ถ้ามีปริมาณของเอทธิลีนมากในส่วนของรากจะเกิดการกระตุ้นให้มีการเพิ่มระดับของเอทธิลีนที่ยอดด้วย ซึ่งกลไกการกระตุ้นนี้ยังไม่เข้าใจเด่นชัดนัก เอทธิลีนอาจจะเคลื่อนที่ผ่านพืชในรูปของ ACC

114
ความสัมพันธ์ของโครงสร้างและคุณสมบัติของเอทธิลีน
คุณสมบัติของเอทธิลีนจะขึ้นอยู่กับลักษณะต่อไปนี้ 1. มีแขนแบบ double bond และไม่อิ่มตัว ซึ่งจะมีคุณสมบัติมากกว่า single หรือ triple bond 2. คุณสมบัติจะลดลงเมื่อโมเลกุลยาวขึ้น 3. แขนแบบ double bond จะต้องอยู่ติดกับคาร์บอนอะตอมสุดท้าย 4. คาร์บอนอะตอมสุดท้ายต้องไม่มีประจุบวก

115
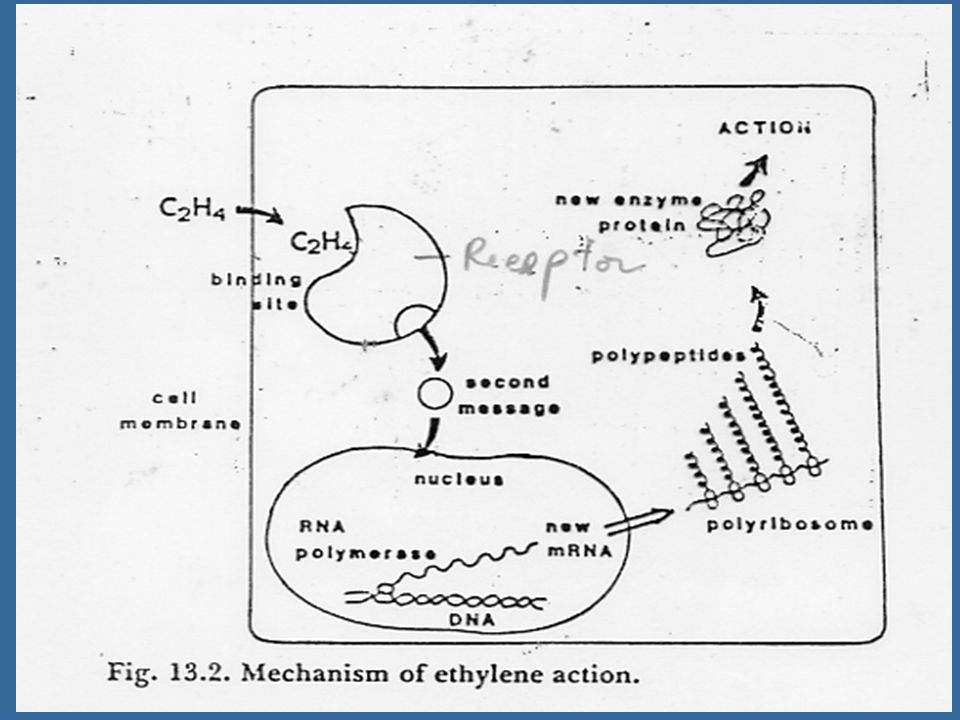
กลไกการทำงานของเอทธิลีน
เอทธิลีนจะก่อให้เกิดผลต่อการเจริญเติบโตของพืช โดยการรวมกับ receptor site ซึ่งมีโลหะรวมอยู่ในโมเลกุลด้วย และคาดว่าโลหะดังกล่าวคือทองแดงและ analogues ของเอทธิลีนจะสามารถแข่งขันเข้ารวมกับ receptor site และก่อให้เกิดผลคล้ายคลึงกับผลของเอทธิลีน บริเวณที่เอทธิลีนรวมกับ receptor site

118
กลไกการทำงานของเอทธิลีน
เอทธิลีนจะควบคุม RNA ให้สังเคราะห์โปรตีน มีเอนไซม์หลายชนิดมีกิจกรรมเพิ่มขึ้นหลังจากที่ได้รับเอทธิลีนแล้ว เช่น เซลลูเลส (Cellulase) เพอร์ออกซิเดส (Peroxidase) ฟีนีลอะลานีน แอมโมเนีย ไลเอส (Phenylalanine ammonia lyase) และฟอสฟาเตส (Phosphatase)
 เพอร์ออกซิเดส (Peroxidase) ฟีนีลอะลานีน แอมโมเนีย ไลเอส (Phenylalanine ammonia lyase) และฟอสฟาเตส (Phosphatase)")
119
กลไกการทำงานของเอทธิลีน
ผลของเอทธิลีนที่เกิดอย่างรวดเร็วนั้น จะไม่สามารถหยุดยั้งด้วย สารระงับการสร้าง RNA หรือโปรตีน เช่น แอคติโนมัยซิน ดี และไซโคเฮกซิไมด์ การที่เอทธิลีนละลายได้ดีมากในไขมัน จึงคาดว่า receptor site ก็อยู่ร่วมกับเยื่อหุ้มเซลล์

120
ผลของเอทธิลีนต่อพืช 1. กระตุ้นให้ผลไม้สุก
2. กระตุ้นการเปลี่ยนแปลงทางคุณภาพ เช่น กระตุ้นให้เกิด Abcission zone ขึ้น 3. กระตุ้นให้พืชออกจากการพักตัว 4. กระตุ้นให้เกิดดอกตัวเมียมากขึ้นในพืช Dioecious

121
การใช้ Hormone และ Growth Regulation บางชนิด
ไม้ผล Paclobutrazol ชักนำการออกดอก Ethephon ออกดอก เปลี่ยนเพศการสุกของผลไม้ NAA เปลี่ยนเพศ ลดการร่วงของดอกและผล GA ยืดช่อ เพิ่มขนาดของผล IBA เร่งราก สับปะรด Ethepon เร่งการออกดอก การสุกสม่ำเสมอ ยางพารา เร่งน้ำยาง พืชอื่นๆ เร่งการเลื้อยของเถา ลดการร่วงของดอกและผล เพิ่มขนาด

122
The End

งานนำเสนอที่คล้ายกัน









 คริษฐา เสมานิตย์.>")
















