ดาวน์โหลดงานนำเสนอ
1
การตอบสนองต่อความยาวของวัน
รองศาสตราจารย์ ดร. ดนัย บุณยเกียรติ ภาควิชาพืชสวน คณะเกษตรศาสตร์ มหาวิทยาลัยเชียงใหม่

2
การออกดอกของพืชคือการเปลี่ยนแปลงของการเจริญเติบโตทางด้านกิ่งก้านสาขา (Vegetative Growth) สู่การเจริญทางด้านการสืบพันธุ์ (Reproductive Growth) เพราะดอกก็คืออวัยวะสืบพันธุ์ของต้นไม้นั่นเอง หลังจากที่พืชมีการเจริญเติบโตทางด้านกิ่งก้านสาขาจนถึงอายุที่มีความพร้อมที่จะออกดอก (Ripeness to Flower) ก็จะเกิดการเปลี่ยนแปลงขึ้นที่ใบพืช ซึ่งจะส่งผลให้เกิดการออกดอกได้
 สู่การเจริญทางด้านการสืบพันธุ์ (Reproductive Growth) เพราะดอกก็คืออวัยวะสืบพันธุ์ของต้นไม้นั่นเอง หลังจากที่พืชมีการเจริญเติบโตทางด้านกิ่งก้านสาขาจนถึงอายุที่มีความพร้อมที่จะออกดอก (Ripeness to Flower) ก็จะเกิดการเปลี่ยนแปลงขึ้นที่ใบพืช ซึ่งจะส่งผลให้เกิดการออกดอกได้")
3
ปัจจัยซึ่งสำคัญที่สุดที่มีผลกระทบต่อการออกดอก คือ แสง โดยที่แสงมีผลต่อการออกดอกทั้งในแง่ของช่วงเวลาที่ได้รับแสง (Photoperiod) คุณภาพของแสง (Wave length) และ Irradiance หรือ Radiant energy ทั้ง 3 ส่วนของแสงมักจะมีผลกระทบต่อการออกดอกอย่างมีปฏิสัมพันธ์กัน (Interaction)
 คุณภาพของแสง (Wave length) และ Irradiance หรือ Radiant energy ทั้ง 3 ส่วนของแสงมักจะมีผลกระทบต่อการออกดอกอย่างมีปฏิสัมพันธ์กัน (Interaction)")
5
Photoperiodism คือ การตอบสนองของพืชต่อความยาวของวัน โดยเฉพาะความสามารถใน การออกดอก พืชจำนวนมากที่จะออกดอกได้เมื่อมีความยาวของวันที่เหมาะสม การค้นพบเรื่อง Photoperiodism นี้เกิดจากการศึกษาของ W.W.Garner และ H.A.Allard (1923) ซึ่งทดลองปลูกยาสูบพันธุ์ Maryland Mammoth ซึ่งจะออกดอกในฤดูหนาวโดยปลูกในเรือนกระจกเท่านั้น นอกจากนั้นเขายังได้ศึกษาถึงการออกดอกของ ถั่วเหลืองทำให้สามารถแบ่งพืชออกเป็น 3 กลุ่ม คือ
 ซึ่งทดลองปลูกยาสูบพันธุ์ Maryland Mammoth ซึ่งจะออกดอกในฤดูหนาวโดยปลูกในเรือนกระจกเท่านั้น นอกจากนั้นเขายังได้ศึกษาถึงการออกดอกของ ถั่วเหลืองทำให้สามารถแบ่งพืชออกเป็น 3 กลุ่ม คือ")
6
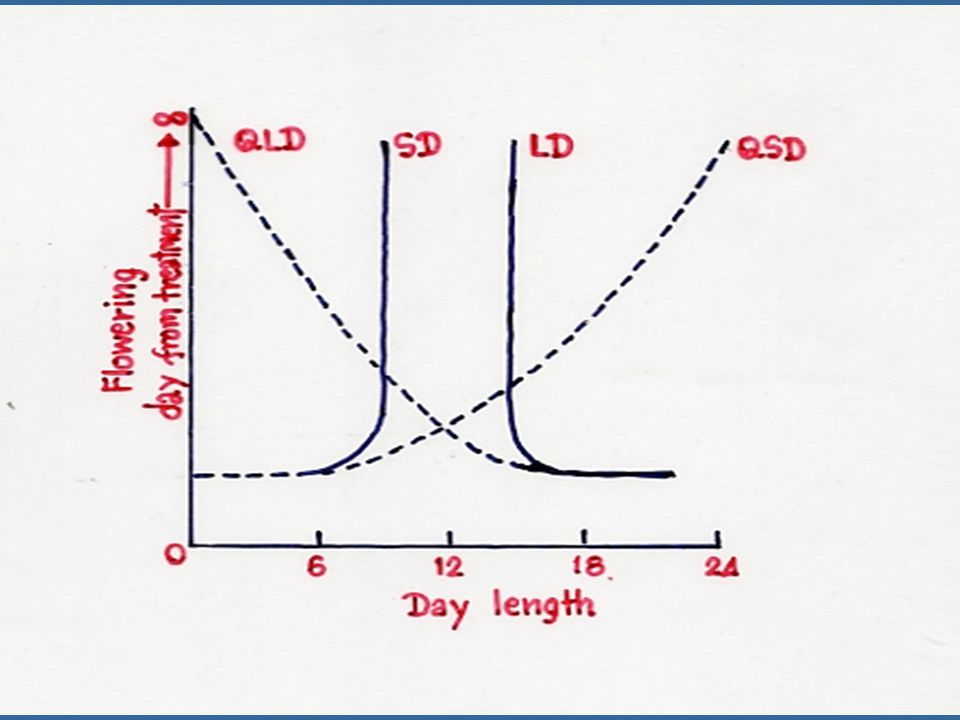
1. พืชที่จะออกดอกเมื่อได้รับช่วงวันสั้น เรียกว่า พืชวันสั้น (Short Day Plant หรือ SDP) พืชชนิดนี้จะออกดอกได้เมื่อได้รับความยาวของวันสั้นกว่าวันวิกฤต (Critical day length) 2. พืชที่จะออกดอกเมื่อได้รับช่วงวันยาว เรียกว่า พืชวันยาว (Long Day Plant หรือ LDP) จะออกดอกเมื่อได้รับความยาวของวันซึ่งยาวกว่า วันวิกฤต (Critical day length) 3. พืชที่ไม่ตอบสนองต่อความยาวของวันเรียกว่า Day neutral plant
 จะออกดอกเมื่อได้รับความยาวของวันซึ่งยาวกว่า วันวิกฤต (Critical day length) 3. พืชที่ไม่ตอบสนองต่อความยาวของวันเรียกว่า Day neutral plant.")
7
พืชบางชนิดจะไม่สามารถออกดอกได้เลย ถ้าหากว่าได้รับความยาวของวันที่ไม่เหมาะสม ซึ่งจัดเป็นพืชพวก Obligate Photoperiodic Plant ซึ่งมีทั้งพืชวันสั้น เช่น Xanthium pennsylvanicum และวันยาว เช่น Hyoscyamus niger พืชบางชนิดจะออกดอกได้เร็วเมื่ออยู่ในสภาพวันสั้นหรือยาว แต่ถ้าไม่ได้รับความยาวของวันตามต้องการก็จะสามารถออกดอกได้เช่นกัน แต่ต้องใช้เวลานานขึ้น พืชพวกนี้เป็น Quantitative Photoperiodic Plant ซึ่งได้แก่ พวกต้องการวันสั้นคือ Salvia splendens ข้าวและฝ้าย ส่วนพืชวันยาวในกลุ่มนี้คือ ข้าวสาลี และ Flax พืชที่เป็น Obligate Photoperiodic Plants จะมีความยาวของวันวิกฤต (Critical Day Length) ที่แน่นอน
 ที่แน่นอน")
8
ตัวอย่างของพืชวันสั้นและวันยาวบางชนิด
1. Qualitative Short Day Amaranthus caudatus Ipomoea hederacea (Morning Glory) Chenopodium album (Pigweed) Nicotiana tabacum Chrysanthemum moriforlium (สายพันธุ์ Maryland Mammoth) Coffea arabica (กาแฟ) Xanthium strumarium (Cocklebur) Euphorbia pulcherrima (Poinsettia) Fragaria (สตรอเบอรี่) Glycine max (ถั่วเหลือง)
 Chenopodium album (Pigweed) Nicotiana tabacum. Chrysanthemum moriforlium (สายพันธุ์ Maryland Mammoth) Coffea arabica (กาแฟ) Xanthium strumarium (Cocklebur) Euphorbia pulcherrima (Poinsettia) Fragaria (สตรอเบอรี่) Glycine max (ถั่วเหลือง)")
9
พืชวันสั้น 2. Quantative Short Day Canabis sativa (Hemp)
Cosmos bipinnatus (Cosmos) Gossypium hirsutum (ฝ้าย) Oryza sativa (ข้าว) Saccharum offinarum (อ้อย) Salvia splendens
 Gossypium hirsutum (ฝ้าย) Oryza sativa (ข้าว) Saccharum offinarum (อ้อย) Salvia splendens.")
10
พืชวันยาว 1. Qualitative Long Day Anethum graeolens (Dill) Avena sativa (ข้าวโอ๊ต) Dianthus superbus (คาร์เนชั่น) Mentha piperita (peppermint) Raphanus sativus (แรดิช) Spinacia oleracea (ปวยเหล็ง) Trifolium spp.(Clover)
 Raphanus sativus (แรดิช) Spinacia oleracea (ปวยเหล็ง) Trifolium spp.(Clover)")
11
พืชวันยาว 2. Quantitative Long Day Anthirrhinum majus (ลิ้นมังกร) Beta vulgaris (หัวบีท) Brassica rapa (เทอร์นิพ) Lactuca sativa (ผักสลัด) Petunia hybrida (พิทูเนีย) Pisum sativum(ถั่วลันเตา) Triticum aestivum (ข้าวสาลีฤดูใบไม้ผลิ)
 Petunia hybrida (พิทูเนีย) Pisum sativum(ถั่วลันเตา) Triticum aestivum (ข้าวสาลีฤดูใบไม้ผลิ)")
12
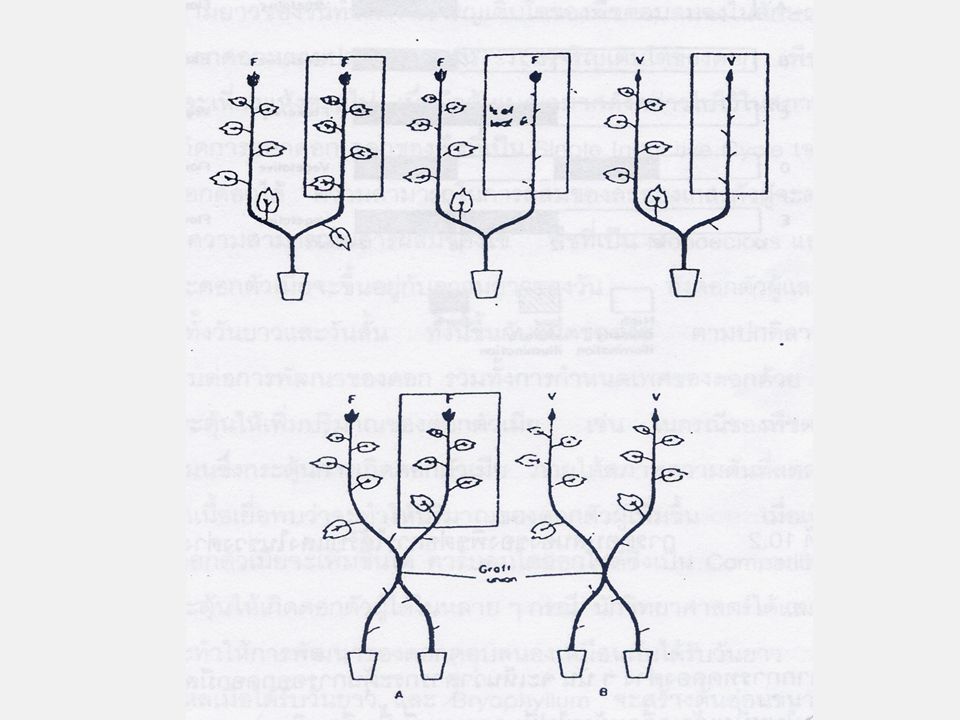
พืชบางกลุ่มที่ต้องการความยาวของวันทั้ง 2 ชนิด เช่น พืชบางชนิดต้องการวันยาวก่อนแล้วตามด้วยวันสั้นเพื่อออกดอก พืชพวกนี้เรียกว่า พืชวันยาวตามด้วยวันสั้น (Long-Short Day Plant) ส่วนพืชที่ต้องการวันสั้นก่อนแล้วตามด้วยวันยาว เรียกว่า พืชวันสั้นตามด้วยวันยาว (Short-Long Day Plant)
 ส่วนพืชที่ต้องการวันสั้นก่อนแล้วตามด้วยวันยาว เรียกว่า พืชวันสั้นตามด้วยวันยาว (Short-Long Day Plant)")
14
ตัวอย่างพืชซึ่งจัดเป็นพืชวันยาวตามด้วยวันสั้น คือ Bryophyllum และ Crassulcean spp. พืชพวกนี้ในเขตอบอุ่นจะออกดอกตอนปลายฤดูร้อนหรือฤดูใบไม้ร่วง แต่จะไม่ออกดอกในฤดูใบไม้ผลิ ตัวอย่างพืชพวกวันสั้นตามด้วยวันยาว คือ Trifolium repens ซึ่งจะออกดอกตอนปลายฤดูใบไม้หรือต้นฤดูร้อน

15
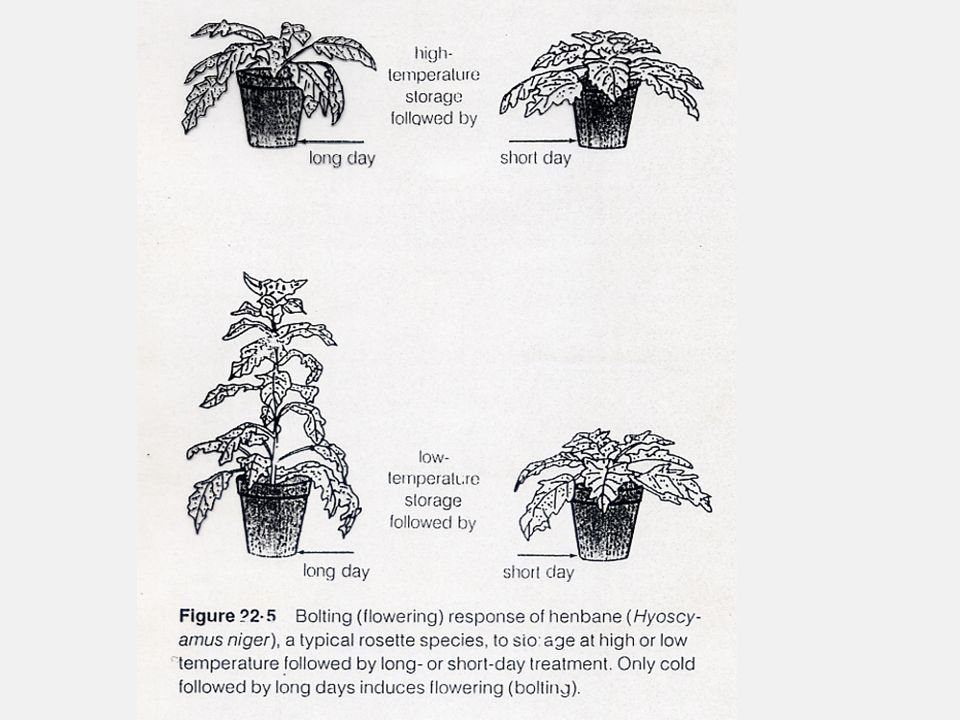
พืชที่ต้องการอุณหภูมิต่ำแล้วตามด้วยวันยาว คือ พืชในกลุ่ม Biennial เช่น พืชตระกูลกะหล่ำ หัวบีท และธัญพืชเมืองหนาว(Winner grain) พืชวันยาวที่ต้องการอุณหภูมิสูง คือ ปวยเหล็ง พืชวันสั้นที่ต้องการอุณหภูมิต่ำเพื่อออกดอก คือ เบญจมาศ พืชวันสั้นที่ต้องการอุณหภูมิสูงเพื่อการออกดอก คือ China Aster

16
การสร้างการ “ไหล” ของสตรอเบอรี่จะเกิดขึ้นภายใต้สภาพวันยาว
การสร้างหัวของเจรูซาเร็ม อาร์ติโช๊ค และมันฝรั่งพันธุ์ป่าจะเกิดเมื่อได้รับวันสั้น หอมหัวใหญ่นั้นจะสร้างหัวเมื่อได้รับวันยาว พืชจะออกดอกเนื่องจากการตอบสนองต่อสภาพแวดล้อม เช่น ความยาวของวันและอุณหภูมิ ใบจะต้องเข้าสู่สภาพที่เรียกว่า Ripeness to respond และเนื้อเยื่อเจริญต้องมีศักยภาพที่ตอบสนองต่อการกระตุ้น (Stimulus) จากใบ
 จากใบ.")
17
ความไวต่อการตอบสนองต่อความยาวของวันจะผันแปรไปโดยสามารถแสดงจำนวน Photoperiodic Cycles
Xanthium ต้องการช่วงได้รับวันสั้นเพียง 1 รอบเท่านั้นก็สามารถออกดอกได้ ถั่วเหลืองถ้าได้รับวันสั้นหลายรอบจำนวนดอกจะเพิ่มมากขึ้น พืชที่ต้องการรอบของช่วงแสง 1 รอบนี้ เรียกว่า Single Inductive Cycle เช่น Cocklebur หรือ Xanthium และ Japanese Morning Glory พืชวันยาวที่เป็น Single Inductive Cycle เช่น Dill Rape และข้าวบาร์เลย์

18
Karl Hamner และ James Bonner
ทดลองให้รอบของกลางวันและกลางคืนหลายแบบซึ่งเมื่อเวลากลางวันกลางคืนรวมกันแล้วไม่เท่ากับ 24 ชั่วโมง พบว่า ความมืดวิกฤต (Critical night) จะคงที่ตลอดทุกการทดลอง แต่วันวิกฤต (Critical day) ไม่คงที่ แสดงให้เห็นว่า ความมืดมีความสำคัญต่อการออกดอก ในอีกการทดลองหนึ่งใช้ การให้ความมืดในช่วงกลางวันแสงและให้แสงในช่วงกลางคืน การให้ความมืดในช่วงกลางวันมีผลกระทบต่อการออกดอกน้อยมากหรือไม่มีเลย แต่การให้แสงในช่วงกลางคืนสามารถระงับการออกดอกของ Xanthium ซึ่งเป็นพืชวันสั้นได้ ต่อมาพบว่าการกระทำดังกล่าวยังกระตุ้นให้พืชวันยาวออกดอกได้
 จะคงที่ตลอดทุกการทดลอง แต่วันวิกฤต (Critical day) ไม่คงที่ แสดงให้เห็นว่า ความมืดมีความสำคัญต่อการออกดอก. ในอีกการทดลองหนึ่งใช้ การให้ความมืดในช่วงกลางวันแสงและให้แสงในช่วงกลางคืน การให้ความมืดในช่วงกลางวันมีผลกระทบต่อการออกดอกน้อยมากหรือไม่มีเลย แต่การให้แสงในช่วงกลางคืนสามารถระงับการออกดอกของ Xanthium ซึ่งเป็นพืชวันสั้นได้ ต่อมาพบว่าการกระทำดังกล่าวยังกระตุ้นให้พืชวันยาวออกดอกได้")
19
Karl Hamner และ James Bonner
จากการทดลองนำไปสู่การค้นพบปรากฏการณ์ Night break การให้แสงสีแดงแล้วตามด้วยแสง Far red แก่ต้น Xanthium ที่ตรงกึ่งกลางของช่วงมืด พบว่า ถ้าให้ Far red ทันทีหลังจากให้แสงสีแดงต้น Xanthium สามารถออกดอกได้ แต่ถ้าให้แสง Far red ช้าไปอีก 30 นาที ต้น Xanthium ไม่ออกดอก ดังนั้น Pfr สามารถระงับกระบวนการกระตุ้น การออกดอกของใบได้ภายใน 30 นาที

20
Karl Hamner และ James Bonner
ถั่วเหลืองจะออกดอกเมื่อได้รับ Inductive Cycle 3 รอบ โดยแต่ละรอบได้รับแสง 12 ชั่วโมง และความมืด 12 ชั่วโมง แต่ถ้าให้แสง 36 ชั่วโมง และความมืด 36 ชั่วโมง ถั่วเหลืองจะไม่ออกดอก แม้จำนวนชั่วโมงที่ได้รับแสงรวมจะเท่ากันก็ตาม

21
Karl Hamner และ James Bonner
Xanthium ซึ่งเป็นพืชวันสั้น ซึ่งมี Critical day length เท่ากับ 15.5 ชั่วโมง ดังนั้นจึงเหลือช่วงมืด 8.5 ชั่วโมง นั่นคือ การออกดอกจะเกิดขึ้นเมื่อได้รับช่วงมืดยาวเท่ากับ 8.5 ชั่วโมงหรือมากกว่า ถ้าหาก Xanthium ได้รับแสง 4 ชั่วโมงและช่วงมืด 8 ชั่วโมง Xanthium จะไม่ออกดอก แม้ว่าช่วงความยาววันจะน้อยกว่า Critical day length ก็ตาม แต่ Xanthium จะออกดอกได้ เมื่อได้รับความยาวของวันเท่ากับ 16 ชั่วโมง และได้รับความมืด 32 ชั่วโมง ซึ่งช่วงความยาววันยาวกว่า Critical day length

22
Karl Hamner และ James Bonner
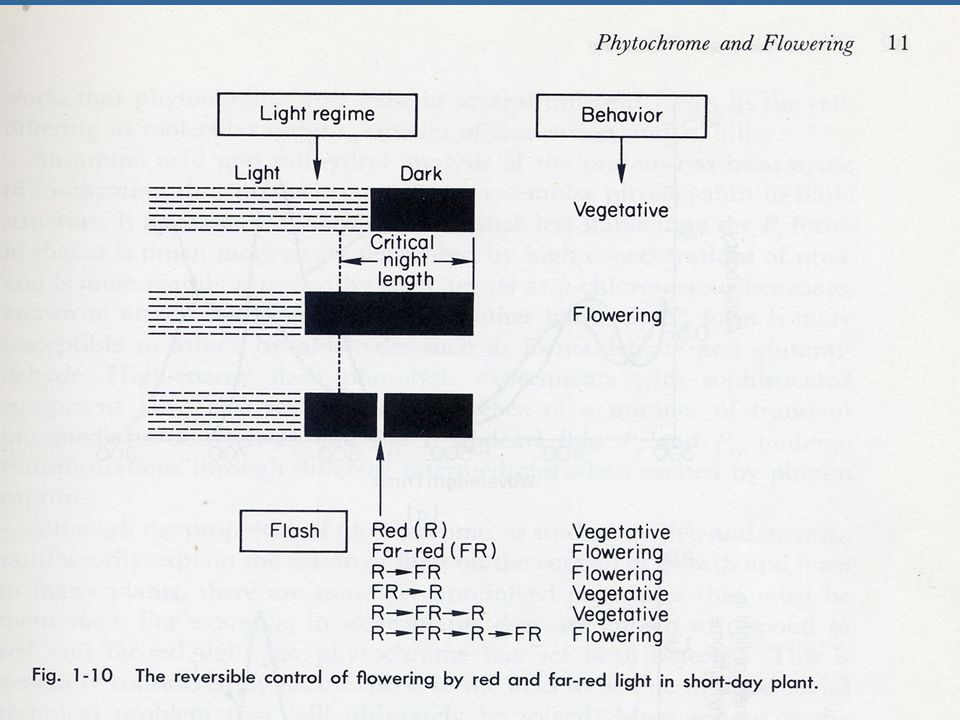
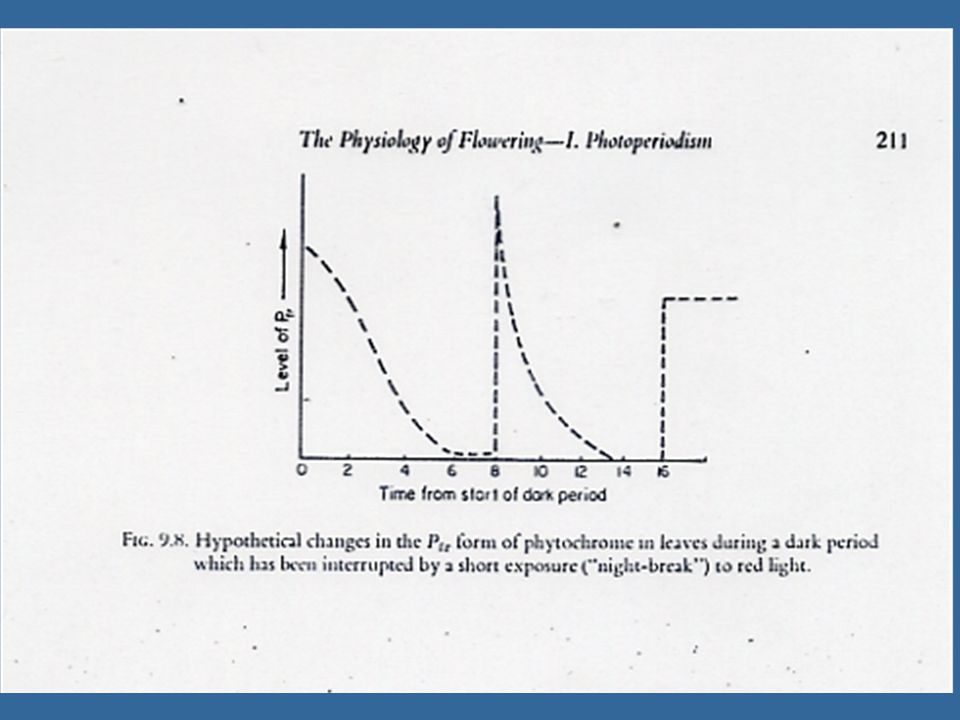
ความสำคัญต่อการออกดอกของพืชวันสั้นเป็นช่วงมืด ดังนั้นพืชวันสั้นจึงมี Critical Dark Period ซึ่งเป็นช่วงมืดที่สั้นที่สุดที่พืชวันสั้นจะออกดอกได้ การให้แสงในช่วงมืดเป็นระยะเวลาสั้นๆ โดยอาจจะสั้นเพียง 1-2 วินาทีนี้เรียกว่า Night Break ซึ่งสามารถลบล้างความสามารถในการออกดอกของพืชวันสั้นและวิธีการนี้จะเกี่ยวข้องกับระบบไฟโตโครม

25
ผลของการให้แสงสีแดงและ Far red ในช่วงมืดต่อการออกดอกของ Xanthium และถั่วเหลือง
กรให้แสง จำนวนดอกของ Xanthium จำนวนดอกของถั่วเหลือง มืดตลอด 6.0 4.0 R 0.0 R,FR 5.6 1.6 R, FR, R, R, FR, R, FR 4.2 1.0 R, FR, R, FR, R - R, FR, R, FR, R, FR 2.4 0.6 หมายเหตุ R คือ แสงสีแดง, FR คือ แสง Far Red

26
พืชวันยาวนั้นค่อนข้างมีกลไกลที่ซับซ้อนกว่าพืชวันสั้นและจะตอบสนองน้อยกว่าและเป็นไปในทางปริมาณต่อ Night break เมื่อเปรียบเทียบกับพืชวันสั้น การออกดอกเนื่องจาก Night break ของ Xanthium เกิดขึ้นเมื่อได้รับแสงเพียง 1-3 วินาที ในชั่วโมงที่8 ยิ่งกว่าไปนั้นขณะที่แสงสีแดงมีประสิทธิภาพสูงสุดในการทำ Night break กับพืชวันสั้น แต่การใช้แสงสีแดงและแสง Far red ผสมกันจะมีประสิทธิภาพในกระตุ้นการออกดอกมากกว่าแสงสีแดง

27
พืชวันสั้นต้องการ Pfr ระดับหนึ่งที่เวลาใดเวลาหนึ่งระหว่างช่วงมืดหรือช่วงมีแสง
ไฟโตโครมนั้นเกิดเป็น 2 ชนิดในพืชวันสั้น ชนิดหนึ่งจำเป็นต่อการกระตุ้นการออกดอกของพืชวันสั้น และจะคงตัว (Stable) ในความมืด อีกชนิดหนึ่งจะไม่คงตัวในความมืด แต่จะระงับการออกดอกของพืชวันสั้นได้ เมื่อปรากฏในช่วงเวลาที่พอเหมาะและ Pfr นั้นน่าจะมีอยู่ 2 ชนิด
 ในความมืด อีกชนิดหนึ่งจะไม่คงตัวในความมืด แต่จะระงับการออกดอกของพืชวันสั้นได้ เมื่อปรากฏในช่วงเวลาที่พอเหมาะและ Pfr นั้นน่าจะมีอยู่ 2 ชนิด.")
30
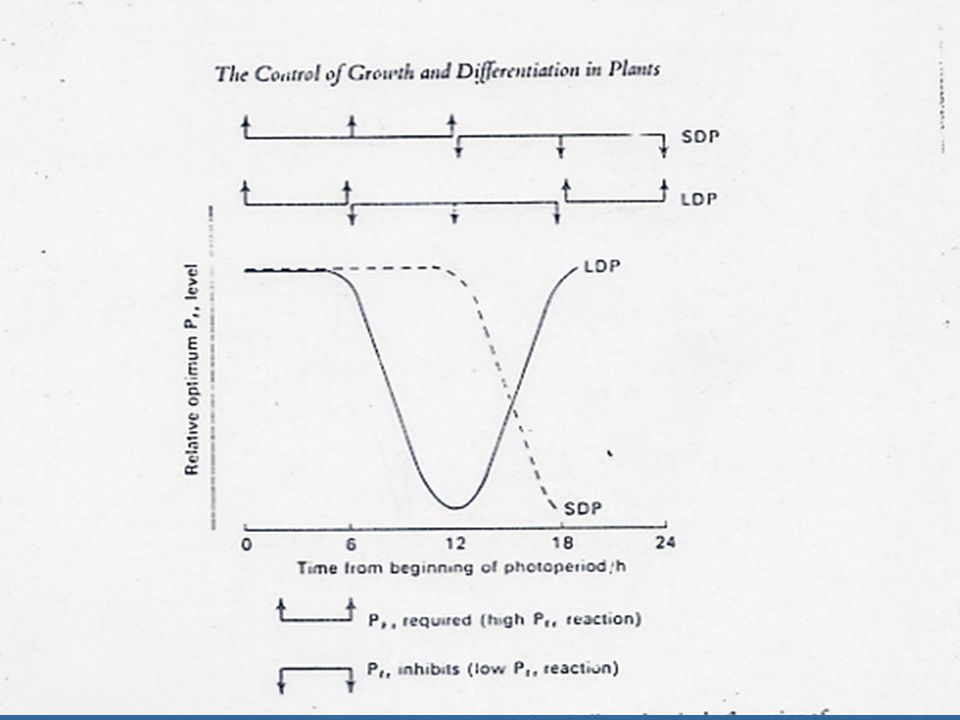
การกระตุ้นการออกดอกของ Pfr จะกลับกันในพืชวันยาว นั่นคือ ระบบไฟโตโครมควบคุมการตอบสนองในการออกดอกของพืชวันยาวและวันสั้นจะสลับกัน คือ ช่วงมืดที่ ยาวจะกระตุ้นให้พืชวันสั้นออกดอก แต่จะระงับการออกดอกของพืชวันยาว ยิ่งไปกว่านั้นพืชทั้งสองชนิดจะตอบสนองต่อ Night Break ซึ่งส่งผลให้เกิด Pfr สูงขึ้น ในช่วงแรกของรอบในการรับแสงพบว่าพืชวันสั้นนั้น การออกดอกจะได้รับการกระตุ้นโดย Pfr สูง ดังนั้นพืชวันสั้นต้องการ Pfr สูงในช่วงแรกของรอบการรับแสงแล้วจึงต้องการ Pfr ต่ำในช่วงต่อมาของรอบการรับแสง

31
การวัดเวลาจากความยาวของวันอาจจะเป็นการวัดช่วงเวลาที่ต้องการสำหรับสารเมตาโบไลท์ จากรูปหนึ่งไปเป็นอีกรูปหนึ่ง ซึ่งจะคล้ายคลึงกับนาฬิกาทราย Xanthium การสร้างสารกระตุ้นการออกดอก (Flowering stimulus) เป็นไปตามระบบของนาฬิกาทราย เป็นที่ชัดเจนว่า Flowering stimulus เพิ่มขึ้นเมื่อได้รับความมืดนานขึ้น
 เป็นไปตามระบบของนาฬิกาทราย. เป็นที่ชัดเจนว่า Flowering stimulus เพิ่มขึ้นเมื่อได้รับความมืดนานขึ้น.")
32
พืชอาจจะวัดความยาวของวันโดย Oscillating circadian clock ซึ่งเหมือนลูกตุ้มนาฬิกาแกว่งไปมา
ถั่วเหลืองซึ่งได้รับ Inductive cycle 7 รอบ แต่ละรอบมีช่วงมืด 64 ชั่วโมง ช่วงมืดนี้ถูกคั่นโดยการให้แสงนานทีละชั่วโมงหลายครั้ง เมื่อศึกษาพืชวันสั้นและวันยาวพบว่ามีการตอบสนองต่อ Night break เป็นจังหวะเหมือนกับถั่วเหลือง คือ ที่เวลาหนึ่งซึ่งแสงระงับการออกดอก การระงับการออกดอกจะเกิดขึ้นอีกใน 24 และ 48 ชั่วโมง ระหว่างเวลาที่ระงับและช่วงของการกระตุ้น ซึ่งชี้ให้เห็นว่าเป็นการวัดเวลาแบบ Oscillating timer

33
ถั่วเหลืองจะออกดอกมากที่สุดเมื่อช่วงแสงและช่วงมืดรวมกันได้ หรือ 72 ชั่วโมง ซึ่งชี้ให้เห็นว่าความมืดไม่ได้ควบคุมการออกดอกทั้งหมด แต่ช่วงแสงก็มีความสำคัญเช่นกัน ส่วนที่รับสัญญาณจากสภาพแวดล้อม คือ ใบ โดยใบจะผลิตสารบางชนิดขึ้นมาเพื่อตอบสนองต่อ Photo Inductive Cycle และสารนี้จะเคลื่อนย้ายไปสู่ยอด Chailachjan เรียกสารที่กระตุ้นให้เกิดการออกดอกที่สร้างที่ใบว่า ฟลอริเจน (Florigen)
")
35
สารกระตุ้นการออกดอกมีสภาพเหมือนฮอร์โมนพืช เพราะเกิดขึ้นในใบหนึ่งแล้วเคลื่อนย้ายไปมีผลกระทบที่เนื้อเยื่อเจริญ ฟลอริเจนจะเป็นความสมดุลของฮอร์โมนหลายชนิดในพืช

36
การตอบสนองต่ออุณหภูมิต่ำ
กลไกของการตอบสนองต่ออุณหภูมิต่ำนั้นอาจจะเกี่ยวข้องกับฮอร์โมนหรือการปิดกั้นกระบวนการ เมตาบอลิสม์ Vernalization คือ การกระตุ้นให้พืชออกดอกโดยการใช้อุณหภูมิต่ำประมาณ 1-7 องศาเซลเซียส โดยเฉพาะกับพืช Bienial และ Perennial พืชพวกนี้ เช่น กะหล่ำปลี เซเลอรี่ หัวบีท และหอมหัวใหญ่ ทั้งนี้รวมกับการกระตุ้นให้ออกดอกเร็วขึ้น โดยได้รับอุณหภูมิต่ำ เช่น ธัญพืชฤดูหนาว (Winter Grain) ผักสลัด และแรดิช เป็นต้น
 ผักสลัด และแรดิช เป็นต้น.")
39
Gassner การศึกษาการออกดอกของธัญพืช เช่น ข้าวสาลี และข้าวไรย์ ซึ่งสามารถแบ่งเป็น 2 กลุ่ม คือ กลุ่มที่หว่านในฤดูใบไม้ร่วงหรือพันธุ์ฤดูหนาว (Winter Varieties) กับพวกที่หว่านในฤดูใบไม้ผลิซึ่งเป็นพันธุ์ฤดูใบไม้ผลิ (Spring Varieties) ทั้ง 2 กลุ่มนี้ แม้ว่าจะหว่านในฤดูต่างกัน แต่จะออกดอกและแก่ในฤดูร้อนถัดไป ถ้าหากหว่านข้าวสาลีฤดูหนาวช้าไปจนถึงฤดูใบไม้ผลิ ข้าวสาลีเหล่านี้จะไม่ออกดอก
 กับพวกที่หว่านในฤดูใบไม้ผลิซึ่งเป็นพันธุ์ฤดูใบไม้ผลิ (Spring Varieties) ทั้ง 2 กลุ่มนี้ แม้ว่าจะหว่านในฤดูต่างกัน แต่จะออกดอกและแก่ในฤดูร้อนถัดไป ถ้าหากหว่านข้าวสาลีฤดูหนาวช้าไปจนถึงฤดูใบไม้ผลิ ข้าวสาลีเหล่านี้จะไม่ออกดอก.")
40
Vernalization ซึ่งโดยเริ่มต้นหมายถึง การกระตุ้น การออกดอกในธัญพืชฤดูหนาวด้วยการได้รับอุณหภูมิต่ำของต้นกล้า เมล็ดชื้นหรือเมล็ดที่กำลังงอก แต่ความหมายในปัจจุบันได้เปลี่ยนไปเป็นการกระตุ้นให้พืชออกดอกโดยการได้รับอุณหภูมิต่ำมาก่อน อายุของพืชซึ่งเหมาะสมต่อการได้รับอุณหภูมิต่ำ พืชบางชนิดจะตอบสนองต่ออุณหภูมิต่ำได้ดีในระยะที่เป็นเมล็ด เช่น ในกรณีของ Winter grain ต่าง ๆ พืชบางชนิดจะตอบสนองต่ออุณหภูมิต่ำได้เมื่อเจริญเติบโตเต็มที่แล้ว เช่น บีท ชนิดตอบสนองต่ออุณหภูมิต่ำได้ในทุกระยะของการเจริญเติบโตโดยที่อายุจะมีระยะที่ตอบสนองดีที่สุดอยู่ระยะหนึ่ง

41
พืชที่ได้รับอุณหภูมิต่ำแล้วสามารถออกดอกได้นั้นยังแบ่งได้เป็น 2 กลุ่ม คือ กลุ่มที่ต้องการอุณหภูมิต่ำเพื่อการออกดอกโดยที่ถ้าไม่ได้รับอุณหภูมิต่ำจะไม่สามารถออกดอกได้ จัดเป็น Obligate requirement for vernalization เช่น กะหล่ำปลี เซเลอรี่ และ Foxglove เป็นต้น พืชบางชนิดหากได้รับอุณหภูมิต่ำจะออกดอกเร็ว แต่ต้นที่ไม่ได้รับอุณหภูมิต่ำก็ออกดอกได้ โดยใช้เวลานานออกไป จัดเป็นกลุ่ม Facultative cold requirement เช่น ผักสลัด และ Pisum sativum บางสายพันธุ์

42
มีพืชหลายชนิดซึ่งมีจุดกำเนิดดอกแล้ว (Flower primordia) แต่การเจริญเติบโตของดอกขึ้นอยู่กับอุณหภูมิที่เหมาะสมซึ่งมักจะเป็นอุณหภูมิต่ำ เช่น ทิวลิป จุดกำเนิดของดอกจะเกิดขึ้นที่อุณหภูมิสูงประมาณ 20 องศาเซลเซียส แต่อุณหภูมิที่จะทำให้ดอกและใบพัฒนาได้ คือ อุณหภูมิ 8-9 องศาเซลเซียส ในตอนแรกและต่อมาต้องการอุณหภูมิ 13 องศาเซลเซียส ซึ่งก็เหมือนกับ ไฮยาซินซ์ (Hyacinth) และแดพโฟดิล (Daffodil) พืชที่แสดงลักษณะการออกดอกแบบนี้ไม่ต้องการ Vernalization เพราะ Vernalization นั้น พืชจะสร้างจุดกำเนิดของดอกหลังจากที่ได้รับอุณหภูมิต่ำแล้วโดยที่อาจจะเกิดที่อุณหภูมิปกติได้
 แต่การเจริญเติบโตของดอกขึ้นอยู่กับอุณหภูมิที่เหมาะสมซึ่งมักจะเป็นอุณหภูมิต่ำ เช่น ทิวลิป จุดกำเนิดของดอกจะเกิดขึ้นที่อุณหภูมิสูงประมาณ 20 องศาเซลเซียส แต่อุณหภูมิที่จะทำให้ดอกและใบพัฒนาได้ คือ อุณหภูมิ 8-9 องศาเซลเซียส ในตอนแรกและต่อมาต้องการอุณหภูมิ 13 องศาเซลเซียส ซึ่งก็เหมือนกับ ไฮยาซินซ์ (Hyacinth) และแดพโฟดิล (Daffodil) พืชที่แสดงลักษณะการออกดอกแบบนี้ไม่ต้องการ Vernalization เพราะ Vernalization นั้น พืชจะสร้างจุดกำเนิดของดอกหลังจากที่ได้รับอุณหภูมิต่ำแล้วโดยที่อาจจะเกิดที่อุณหภูมิปกติได้")
43
ส่วนของพืชที่ตอบสนองต่ออุณหภูมิต่ำจะอยู่ที่ปลายยอดโดยเซลล์ของเนื้อเยื่อเจริญบริเวณนั้นจะตอบสนองต่ออุณหภูมิต่ำ โดยส่วนอื่น ๆ ของต้นอาจจะอยู่ในอุณหภูมิเท่าไรก็ได้ การศึกษาให้อุณหภูมิต่ำบริเวณปลายยอดนี้ได้ผลใน เซเลอรี่ บีท เบญจมาศ และหอมหัวใหญ่

44
II I Intermediat product (B) Precursor(A) End product (C) III D
 Precursor(A) End product (C) III D")
45
สารกระตุ้นการออกดอกที่เกิดจาก Vernalization นี้ Melchers และ Lang ตั้งชื่อว่า เวอนาลิน (Vernalin)
เวอนาลินต่างจากฟลอริเจน ยาสูบพันธุ์ Maryland Mammoth ซึ่งเป็นพืชวันสั้นต่อเชื่อมกับ Hyoscyamus niger ซึ่งเป็นพืชวันยาว และต้องการอุณหภูมิต่ำในการออกดอก พบว่าในสภาพวันยาว Hyoscyamus niger สามารถออกดอกได้โดยไม่ต้องได้รับอุณหภูมิต่ำ แต่ต้นยาสูบเป็นพืชวันสั้นจึงไม่สามารถออกดอกในสภาพวันยาว ดังนั้นจึงแสดงว่า การที่ต้นรับ (Hyoscyamus niger) ออกดอกได้แสดงว่าไม่ได้เกิดจากการถ่ายทอด ฟลอริเจนจากยาสูบ เพราะยาสูบไม่ออกดอก
 ออกดอกได้แสดงว่าไม่ได้เกิดจากการถ่ายทอด ฟลอริเจนจากยาสูบ เพราะยาสูบไม่ออกดอก.")
46
สภาวะที่เกิดผลของอุณหภูมิต่ำ (Vernalized State)
เวอนาลิน ฟลอริเจน หรือ เวอนาลิน ฟลอริเจน การออกดอก พืชที่ต้องการอุณหภูมิต่ำ พืชวันยาวหรือวันสั้นเมื่อ จะต้องได้รับอุณหภูมิต่ำ ได้รับช่วงแสงตามต้องการ พืชอื่นๆ ไม่จำเป็น พืชอื่นๆ ไม่จำเป็นต้องได้รับ ต้องได้รับอุณหภูมิต่ำ ช่วงแสงตามต้องการ

47
Devernalization ซึ่งจะลบล้างด้วยอุณหภูมิสูงและสภาพขาดออกซิเจน ซึ่งพบได้ในเซเลอรี่และธัญพืชบางชนิด ดังนั้นสารที่เกิดขึ้นโดย Vernalization อาจจะไม่เสถียรในอุณหภูมิสูง และในสภาพที่ขาดออกซิเจน

48
ฮอร์โมน คือ สารประกอบอินทรีย์ ซึ่งสามารถมีผลกระทบในปริมาณที่น้อยมาก โดยพืชจะสังเคราะห์ที่ส่วนหนึ่งแล้วเคลื่อนย้ายไปยังอีกส่วนหนึ่ง และมีผลต่อกระบวนการทางสรีรวิทยาที่ค่อนข้างเฉพาะเจาะจง

49
สารควบคุมการเจริญเติบโต (Plant Growth Regulator)เป็นสารเคมีที่สำคัญในการเกษตรเป็นสารอินทรีย์ซึ่งมนุษย์สังเคราะห์ขึ้นมาได้ ซึ่งบางชนิดมีคุณสมบัติเหมือนฮอร์โมนพืช ในบางกรณีนี้ฮอร์โมนจะทดแทนสภาพแวดล้อมที่เหมาะสมได้ ในประเทศไทยการใช้ฮอร์โมนพืชมีวัตถุประสงค์ในทางการเกษตรเพื่อให้มีผลผลิต เพื่อเพิ่มผลผลิตและคุณภาพ และเพื่อความสะดวกในการจัดการฟาร์ม

50
ออกซิน (Auxin) Charles Darwin ซึ่งศึกษาเรื่อง Phototropism ซึ่งพืชจะโค้งงอเข้าหาแสง Darwin ทดลองกับ ต้นกล้าของ Phalaris canariensis และพบว่าโคลีออพไทล์ของพืชชนิดนี้จะตอบสนองต่อการได้รับแสงเพียงด้านเดียวทำให้เกิดการโค้งเข้าหาแสง Darwin สรุปว่าเมื่อต้นกล้าได้รับแสงจะทำให้มี "อิทธิพล" (Influence) บางอย่างส่งผ่านจากส่วนยอดมายังส่วนล่างของโคลีออพไทล์ ทำให้เกิดการโค้งงอเข้าหาแสง ซึ่งนักวิทยาศาสตร์รุ่นต่อมาได้ศึกษาถึง "อิทธิพล" ดังกล่าว
 บางอย่างส่งผ่านจากส่วนยอดมายังส่วนล่างของโคลีออพไทล์ ทำให้เกิดการโค้งงอเข้าหาแสง ซึ่งนักวิทยาศาสตร์รุ่นต่อมาได้ศึกษาถึง อิทธิพล ดังกล่าว.")
51
ออกซิน (Auxin) ต่อมา Boysen-Jensen และ Paal ได้ศึกษาและแสดงให้เห็นว่า "อิทธิพล" ดังกล่าวนี้มีสภาพเป็นสารเคมี Went ได้ทำงานทดลองและสามารถแยกสารชนิดนี้ออกจากโคลีออพไทล์ได้ โดยตัดส่วนยอดของโคลีออพไทล์ของข้าวโอ๊ตแล้ววางลงบนวุ้นจะทำให้สารเคมีที่กระตุ้นการเจริญเติบโตไหลลงสู่วุ้น เมื่อนำวุ้นไปวางลงที่ด้านหนึ่งของโคลีออพไทล์ที่ไม่มียอดด้านใดด้านหนึ่งจะทำให้โคลีออพไทล์ดังกล่าวโค้งได้

52
ออกซิน (Auxin) สารเคมีดังกล่าวได้รับการตั้งชื่อว่า ออกซิน ซึ่งในปัจจุบันพบในพืชชั้นสูงทั่วๆ ไป และมีความสำคัญต่อการเจริญเติบโตของพืช สังเคราะห์ได้จากส่วนเนื้อเยื่อเจริญของลำต้น ปลายราก ใบอ่อน ดอกและผล และพบมากที่บริเวณเนื้อเยื่อเจริญโคลีออพไทด์และคัพภะ รวมทั้งใบที่กำลังเจริญด้วย

53
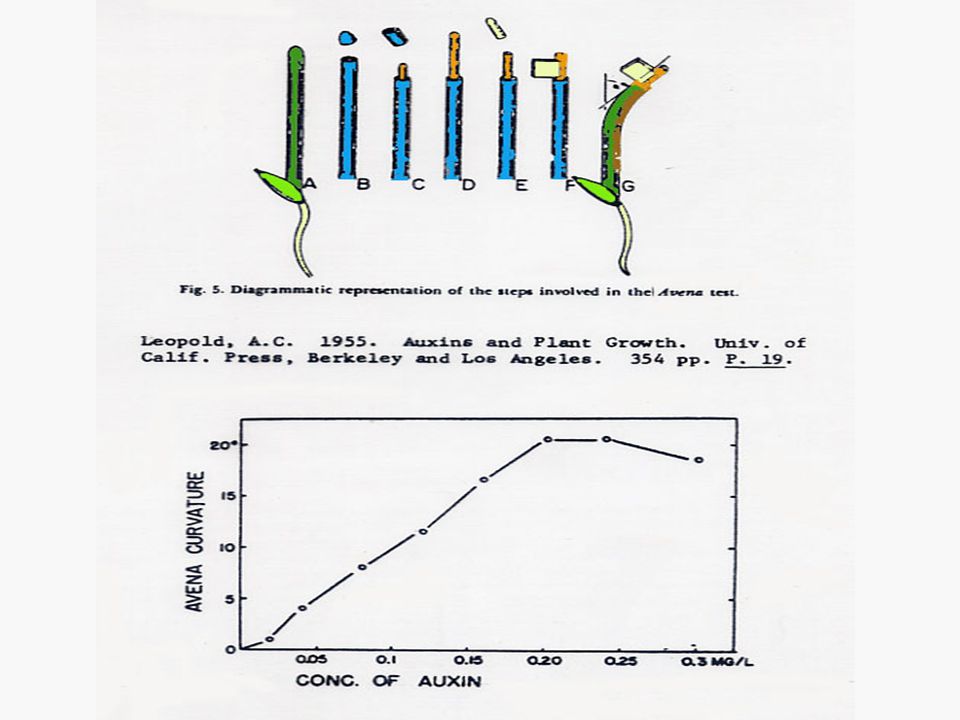
งานทดลองของนักวิทยาศาสตร์ ซึ่งนำมาสู่การค้นพบออกซินในพืช ทุกการทดลอง ทำกับโคลีออพไทด์ของต้นกล้าของพืชตระกูลหญ้า ลูกศร 3 เส้นที่ติดกัน แสดงทิศทางของ unilateral light ในการทดลองของ Went แสดงถึง Went Avena curvature test

54
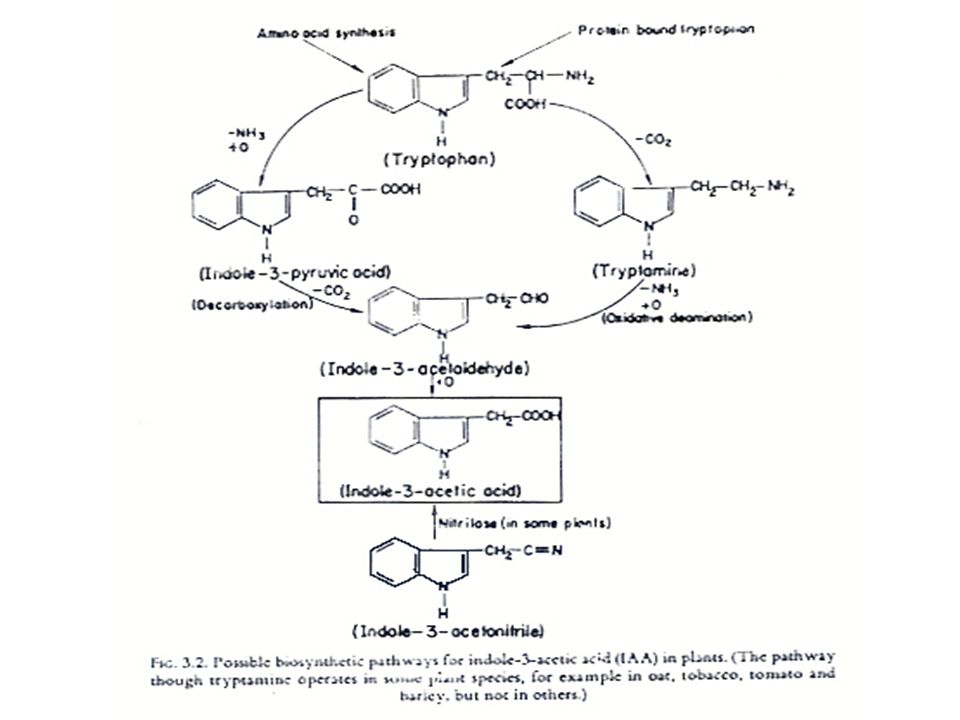
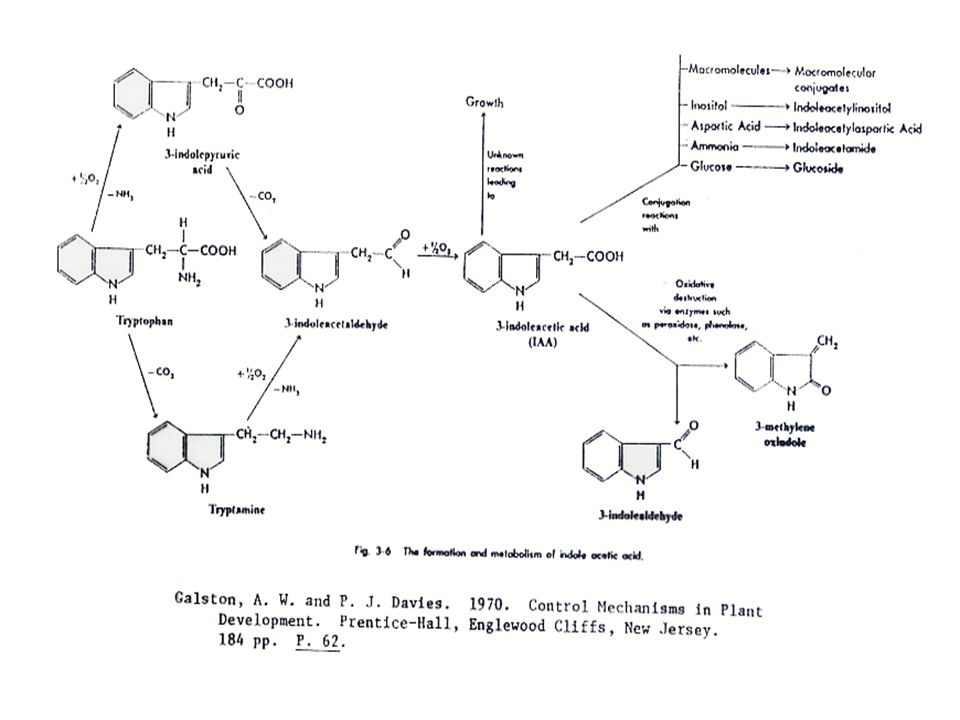
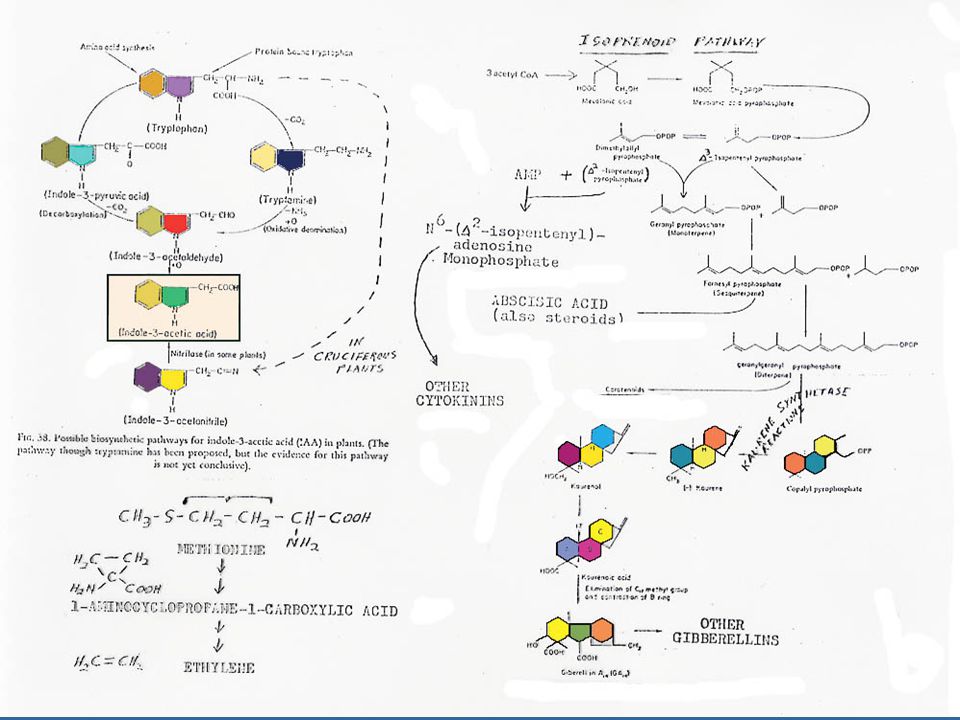
การสังเคราะห์ออกซิน การสังเคราะห์ออกซินนั้นมีกรดอะมิโน L-Tryptophan เป็นสารเริ่มต้น (Precursor) L-Tryptophan เป็นกรดอะมิโนที่มีโครงสร้างของ Indole อยู่ การสังเคราะห์ออกซิน แสดงในรูปที่ 12.2 ซึ่งในการสังเคราะห์ IAA นั้น จะมี IAAld และ IPyA เป็นสารที่พบในระหว่างการสังเคราะห์ ในพืชบางชนิด เช่น ข้าวโอ๊ต ยาสูบ มะเขือเทศ ทานตะวัน และข้าวบาร์เลย์ พบว่า Tryptophan สามารถเปลี่ยนเป็น Tryptamine ได้ ในพืชตระกูลกะหล่ำ Tryptamine อาจจะเปลี่ยนไปเป็น Indoleacetaldoxime แล้วเปลี่ยนไปเป็น IAN แล้วจึงเปลี่ยนเป็น IA
 L-Tryptophan เป็นกรดอะมิโนที่มีโครงสร้างของ Indole อยู่ การสังเคราะห์ออกซิน แสดงในรูปที่ 12.2 ซึ่งในการสังเคราะห์ IAA นั้น จะมี IAAld และ IPyA เป็นสารที่พบในระหว่างการสังเคราะห์ ในพืชบางชนิด เช่น ข้าวโอ๊ต ยาสูบ มะเขือเทศ ทานตะวัน และข้าวบาร์เลย์ พบว่า Tryptophan สามารถเปลี่ยนเป็น Tryptamine ได้ ในพืชตระกูลกะหล่ำ Tryptamine อาจจะเปลี่ยนไปเป็น Indoleacetaldoxime แล้วเปลี่ยนไปเป็น IAN แล้วจึงเปลี่ยนเป็น IA.")
56
การสังเคราะห์ออกซิน การศึกษาเรื่องการสังเคราะห์ออกซินมักศึกษาจากเนื้อเยื่อปลายรากหรือปลายยอด และพบว่า IAA นี้สังเคราะห์ได้ทั้งในส่วนไซโตซอล (Cytosol) ไมโต-คอนเดรีย และคลอโรพลาสต์ ในการศึกษาในปัจจุบันพบว่า Phenylacetic acid หรือ PAA มีคุณสมบัติของออกซินด้วยและสามารถสังเคราะห์ได้จาก L-Phenylalanine โดยพบในคลอโรพลาสต์ และไมโตคอนเดรียของทานตะวัน
 ไมโต-คอนเดรีย และคลอโรพลาสต์ ในการศึกษาในปัจจุบันพบว่า Phenylacetic acid หรือ PAA มีคุณสมบัติของออกซินด้วยและสามารถสังเคราะห์ได้จาก L-Phenylalanine โดยพบในคลอโรพลาสต์ และไมโตคอนเดรียของทานตะวัน.")
57
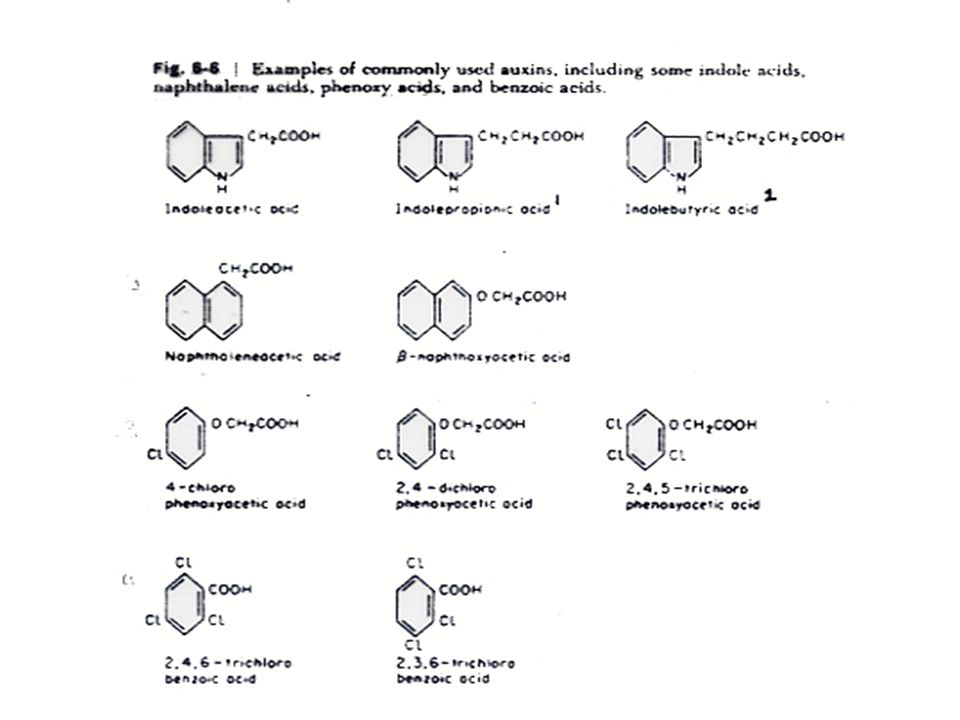
การสังเคราะห์ออกซิน สารสังเคราะห์ที่มีคุณสมบัติของออกซินมีหลายชนิดที่สำคัญทางการเกษตร เช่น สาร 2, 4-dichlorophenoxyacetic acid หรือ 2,4-D ซึ่งใช้ในการกำจัดวัชพืช IBA หรือ Indole butyric acid ใช้ในการเร่งให้ส่วนที่จะนำไปปักชำเกิดรากเร็วขึ้น และ NAA หรือ Napthalene acetic acid จะช่วยในการติดผลของผลไม้บางชนิด

59
โครงสร้างของโมเลกุลที่สำคัญของสารที่จะมีคุณสมบัติของออกซินคือ ต้องประกอบด้วยประจุลบ (Strong Negative Charge) ซึ่งเกิดจากการแตกตัวของกลุ่มคาร์บอกซิล และประจุลบจะต้องอยู่ห่างจากประจุบวก (Weaker Positive Charge) บนวงแหวนด้วยระยะทางประมาณ 5.5 Angstrom
 ซึ่งเกิดจากการแตกตัวของกลุ่มคาร์บอกซิล และประจุลบจะต้องอยู่ห่างจากประจุบวก (Weaker Positive Charge) บนวงแหวนด้วยระยะทางประมาณ 5.5 Angstrom")
60
การสลายตัวของ IAA 1. Photo-oxidation IAA ที่อยู่ในสภาพสารละลายจะสลายตัวได้เมื่อได้รับแสง การเกิด Photo-oxidation ของ IAA จะถูกเร่งโดยการปรากฏของสารสีตามธรรมชาติ หรือที่สังเคราะห์ได้ จึงอาจจะเป็นไปได้ว่าการที่สารสีของพืชดูดซับพลังงานจากแสงแล้วทำให้เกิดการออกซิไดซ์ IAA ซึ่งสารสีที่เกี่ยวข้อง คือ ไรโบฟลา-วิน และไวโอลาแซนธิน (Riboflavin และ Violaxanthin) สารที่เกิดขึ้นเมื่อ IAA สลายตัวโดยแสงคือ 3-methylene-2-oxindole และ Indoleacetaldehyde
 สารที่เกิดขึ้นเมื่อ IAA สลายตัวโดยแสงคือ 3-methylene-2-oxindole และ Indoleacetaldehyde.")
61
2. การออกซิไดซ์โดยเอนไซม์ (Enzymic Oxidation of IAA) พืชหลายชนิดมีเอนไซม์เรียกว่า IAA-oxidase ซึ่งจะคะตะไลท์ สลาย IAA ได้คาร์บอนไดออกไซด์ และเป็นปฏิกิริยาที่ใช้ออกซิเจน IAA-oxidase ผลิตภัณฑ์ที่ได้ คือ 3-methylene-2-oxindole และถูกเมตาโบไลซ์ต่อไป เป็น 3-methyl-2-oxindole
 พืชหลายชนิดมีเอนไซม์เรียกว่า IAA-oxidase ซึ่งจะคะตะไลท์ สลาย IAA ได้คาร์บอนไดออกไซด์ และเป็นปฏิกิริยาที่ใช้ออกซิเจน IAA-oxidase ผลิตภัณฑ์ที่ได้ คือ 3-methylene-2-oxindole และถูกเมตาโบไลซ์ต่อไป เป็น 3-methyl-2-oxindole.")
62
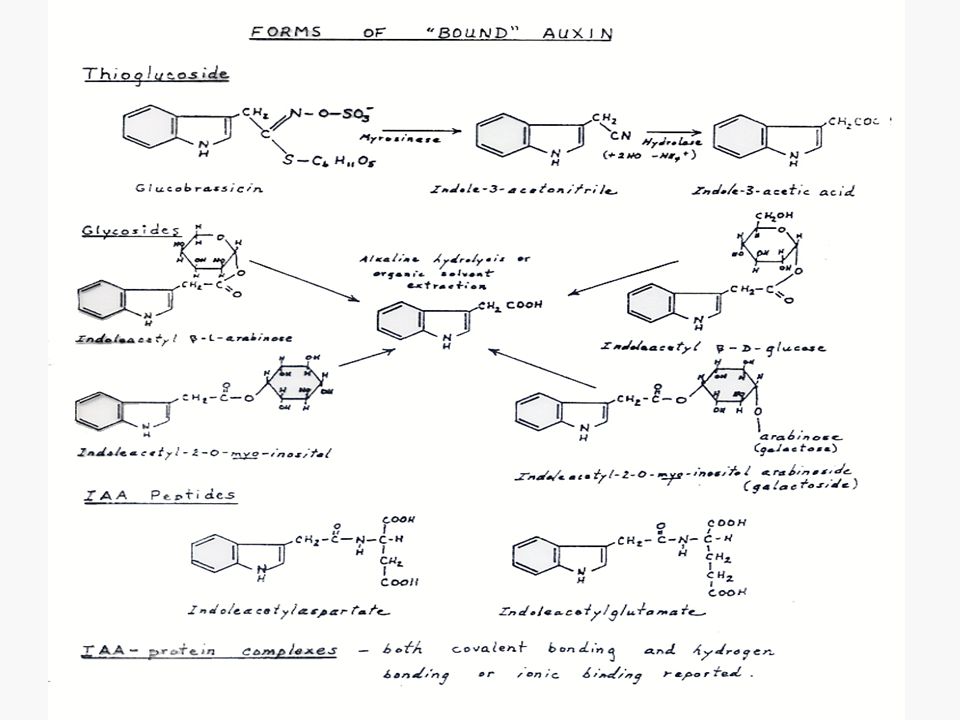
3. รวมกับสารชนิดอื่นในไซโตพลาสต์
4. เปลี่ยนเป็นอนุพันธ์ชนิดอื่น

63
1. Bioassay คือ การวัดปริมาณออกซินโดยใช้ชิ้นส่วนของพืช เช่น โคลีออพไทล์ของข้าวโอ๊ตหรือพืชใบเลี้ยงเดี่ยวอื่นๆ แล้ววัดความโค้งของยอดโดยการวางออกซินที่ต้องการวัดปริมาณลงบนส่วนของโคลีออพไทล์ซึ่งตัดยอดออกแล้ว มุมที่โค้งจะบอกปริมาณของออกซินได้โดยเปรียบเทียบจากเส้นมาตรฐาน (Standard Curve)
.")
67
2. การวัดจากคุณสมบัติทางฟิสิกส์ สกัดจนเป็นสารบริสุทธิ์แล้วจึงใช้เครื่อง Gas Chromatograph ร่วมกับ Mass Spectrometry ในการจำแนกและหาปริมาณ 3. การวัดโดยวิธีเคมีโดยให้ออกซินทำปฏิกิริยากับ Salkowski's Reagent (acidified ferric chloride) หรือใช้ Ehrllch's Reagent ซึ่งจะเกิดสีขึ้นมา จากนั้นวัดความเข้มของสีแล้วเปรียบเทียบกับเส้นมาตรฐาน
 หรือใช้ Ehrllch s Reagent ซึ่งจะเกิดสีขึ้นมา จากนั้นวัดความเข้มของสีแล้วเปรียบเทียบกับเส้นมาตรฐาน.")
68
การเคลื่อนที่ของออกซินในต้นพืช
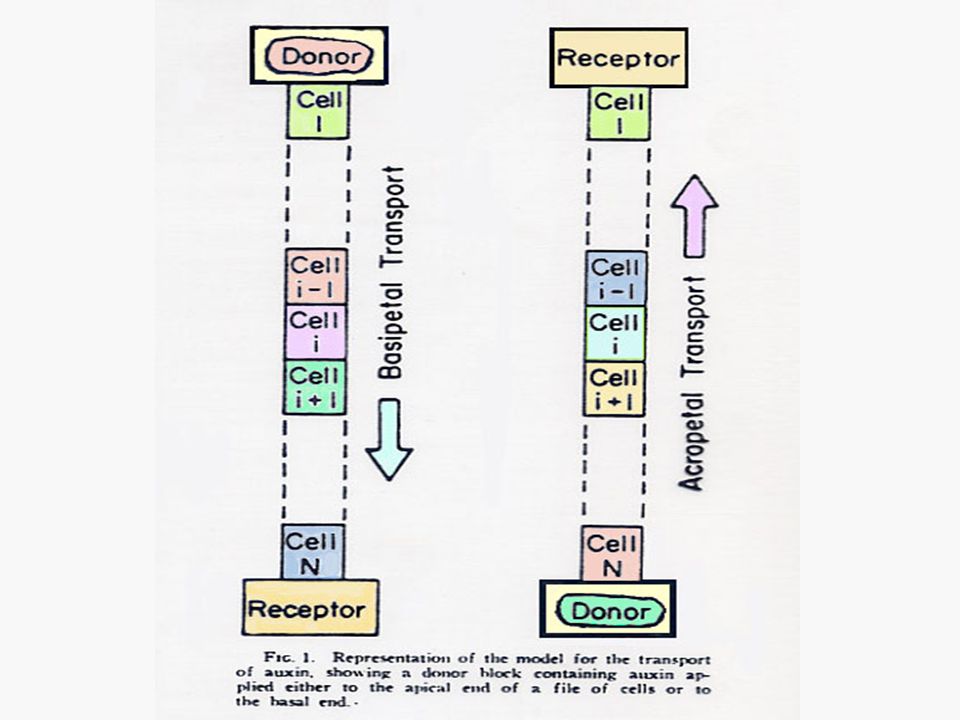
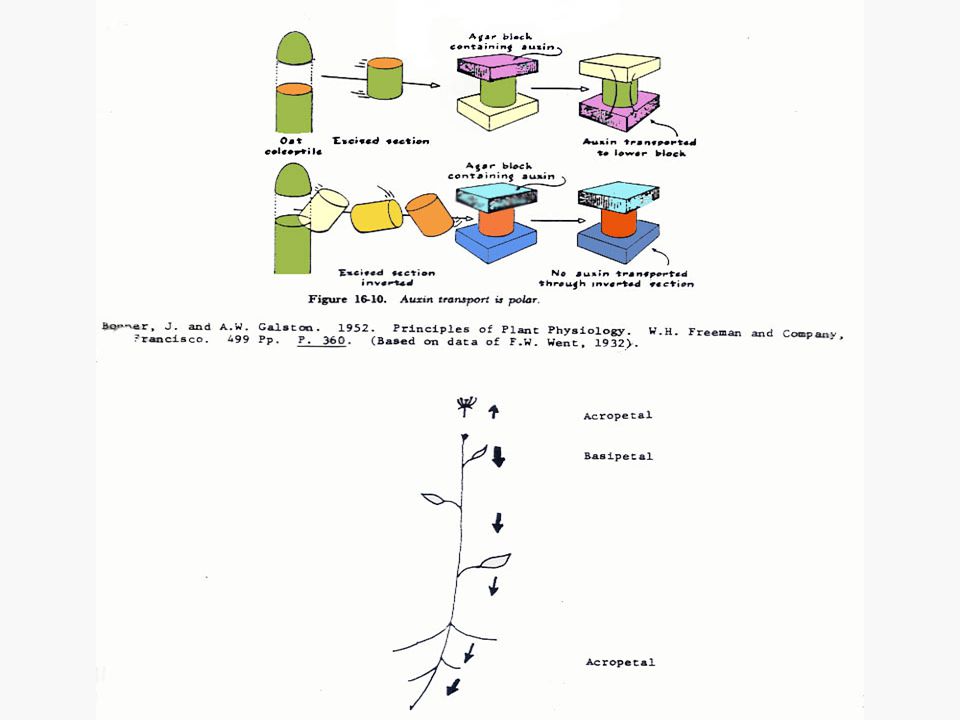
การเคลื่อนที่ของออกซินจะเป็นแบบโพลาไรซ์ (Polarized) คือ เคลื่อนที่ไปตามยาวของลำต้นโดยไปในทิศทางใดทิศทางหนึ่งมากกว่าทิศทางตรงกันข้าม การเคลื่อนที่ของออกซินในส่วนที่อยู่เหนือดิน จะเป็นแบบโพลาร์ เบสิพีตัล (Polar Basipetal) คือ จะเคลื่อนที่จากแหล่งผลิตที่ยอดไปสู่โคนต้น การเคลื่อนที่ของออกซินจะเกิดแบบเบสิพีตัลก็เมื่อท่อนเนื้อเยื่อวางอยู่ในลักษณะปกติของลักษณะทางสัณฐานวิทยาเท่านั้น
 คือ เคลื่อนที่ไปตามยาวของลำต้นโดยไปในทิศทางใดทิศทางหนึ่งมากกว่าทิศทางตรงกันข้าม. การเคลื่อนที่ของออกซินในส่วนที่อยู่เหนือดิน จะเป็นแบบโพลาร์ เบสิพีตัล (Polar Basipetal) คือ จะเคลื่อนที่จากแหล่งผลิตที่ยอดไปสู่โคนต้น การเคลื่อนที่ของออกซินจะเกิดแบบเบสิพีตัลก็เมื่อท่อนเนื้อเยื่อวางอยู่ในลักษณะปกติของลักษณะทางสัณฐานวิทยาเท่านั้น.")
76
ในกรณีของโคลีออพไทล์ของพืชนั้นชี้ให้เห็นว่าออกซินเคลื่อนที่ผ่านเซลล์ทุกเซลล์ลงมาแต่ในกรณีของลำต้นนั้นยังไม่มีหลักฐานชี้ให้เห็นเด่นชัดนัก แต่อาจจะเป็นไปได้ว่าโปรแคมเบียม (Procambium) และแคมเบียม (Cambium) การเคลื่อนที่ของออกซินในรากก็มีลักษณะเป็นโพลาร์ แต่เป็นแบบ อะโครพีตัล

77
กลไกในการทำงานของออกซินในระยะที่ผ่านมาจะมีแนวความคิดเป็นสองอย่าง คือ แนวคิดที่เกี่ยวข้องกับผนังเซลล์เป็นส่วนที่รับผลกระทบของออกซิน และขยายตัว ส่วนอีกแนวคิดหนึ่งมุ่งไปที่ผลของออกซินต่อเมตาบอลิสม์ของกรดนิวคลีอิคในปัจจุบันได้นำสองแนวคิดมาวิเคราะห์ ร่วมกันเพื่อศึกษากลไกในการทำงานของออกซิน และยังศึกษาผลของออกซินต่อเยื่อหุ้มเซลล์ด้วย

78
การขยายตัวของเซลล์จะสัมพันธ์กับการเปลี่ยนแปลงปริมาณและกิจกรรมของเอนไซม์ โดยที่ออกซินจะมีบทบาทต่อ กระบวนการเมตาบอลิสม์ของกรดนิวคลีอิค โดยการศึกษาจากการเพาะเลี้ยงเนื้อเยื่อที่เป็นไส้ของต้นยาสูบ (Tobacco Pith) ซึ่งจะเจริญไปเป็นกลุ่มเนื้อเยื่อ (Callus) นั้นพบว่ามีปริมาณของ RNA เพิ่มมากขึ้น ทั้งนี้เพราะออกซินจะกระตุ้นให้มีการสังเคราะห์ RNA มากขึ้น แล้วส่งผลไปถึงการเจริญของกลุ่มเนื้อเยื่อ ถ้าหากใช้สารระงับการสังเคราะห์โปรตีนหรือ RNA ความสามารถในการกระตุ้นการเจริญเติบโตของออกซินจะหายไป
 ซึ่งจะเจริญไปเป็นกลุ่มเนื้อเยื่อ (Callus) นั้นพบว่ามีปริมาณของ RNA เพิ่มมากขึ้น ทั้งนี้เพราะออกซินจะกระตุ้นให้มีการสังเคราะห์ RNA มากขึ้น แล้วส่งผลไปถึงการเจริญของกลุ่มเนื้อเยื่อ ถ้าหากใช้สารระงับการสังเคราะห์โปรตีนหรือ RNA ความสามารถในการกระตุ้นการเจริญเติบโตของออกซินจะหายไป")
79
ออกซินมีผลต่อระดับเอนไซม์ โดยผ่านทางการสังเคราะห์ RNA นอกจากนั้นออกซินยังมีผลกระทบต่อกิจกรรมของเอนไซม์โดยตรง เช่น การกระตุ้นให้เอนไซม์เกิดกิจกรรมหรือเปลี่ยนรูปมาอยู่ในรูปที่มีกิจกรรมได้ นักวิทยาศาสตร์ได้มุ่งความสนใจไปสู่เอนไซม์ที่สัมพันธ์กับกระบวนการขยายตัวของเซลล์ เซลล์พืชจะมีผนังเซลล์อยู่ข้างนอกสุด ดังนั้นการเจริญของเซลล์จะเกิดขึ้นได้เมื่อคุณสมบัติของผนังเซลล์เปลี่ยนไปในทางที่ก่อให้เกิดการขยายตัวของโปรโตพลาสต์

80
เซลล์พืชทุกชนิดที่ผ่านขั้นตอนของเนื้อเยื่อเจริญมาแล้วจะผ่านขั้นตอนการเจริญเติบโต 2 ขั้น คือ การแบ่งเซลล์และการขยายตัวแวคคิวโอขึ้นภายในเซลล์ (Vacuolation) การขยายตัวของเซลล์นั้น ไม่เพียงแต่ผนังเซลล์ยืดตัวเท่านั้น แต่ยังมีการเพิ่มความหนาของผนังเซลล์เพราะมีสารใหม่ ๆ ไปเกาะด้วย

81
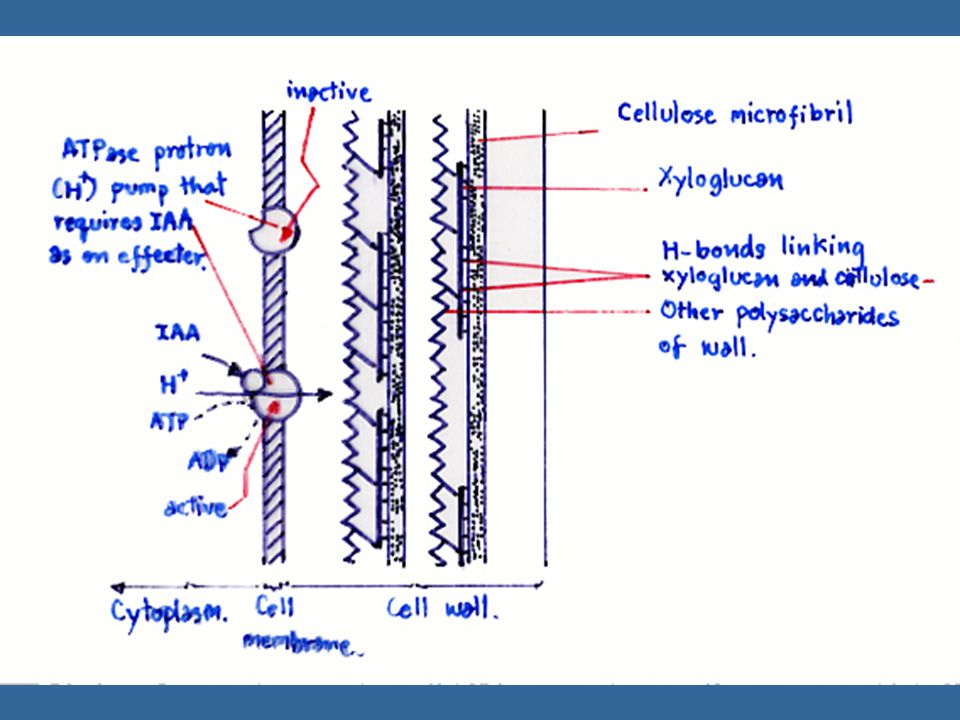
นักวิทยาศาสตร์ได้นำเอาโคลีออพไทล์หรือลำต้นที่ไม่ได้รับแสงไปแช่ลงในสภาพที่มี pH ประมาณ 3 ปรากฏว่าโคลีออพไทล์และลำต้นสามารถยืดตัวได้ และเรียกปรากฏการณ์นี้ว่า "Acid Growth Effect" ซึ่งให้ผลเหมือนกับการให้ออกซินแก่พืช การทดลองนี้ได้นำไปสู่การศึกษาที่แสดงว่าออกซิน กระตุ้นการปลดปล่อย H+ หรือโปรตอนจากเนื้อเยื่อ ทำให้ pH ของผนังเซลล์ต่ำลง ซึ่งการปลดปล่อย H+ นี้ต้องใช้พลังงานจากการหายใจด้วย สมมุติฐานเกี่ยวกับ "Proton-Pump" นี้ คาดว่าเกิดในเยื่อหุ้มเซลล์

82
การเจริญของเซลล์ต้องการ RNA และโปรตีนในช่วงที่เซลล์ยืดตัว เพราะในการยืดตัวของเซลล์นั้นผนังเซลล์ไม่ได้บางลงไป แต่ยังคงหนาเท่าเดิมหรือหนาขึ้น ดังนั้นจึงต้องมีการสร้างผนังเซลล์เพิ่มขึ้นด้วย ในการสร้างผนังเซลล์นั้นต้องใช้เอนไซม์และ RNA pH ต่ำมีผลต่อการเปลี่ยนคุณสมบัติของผนังเซลล์ในแง่ที่ว่า แขนที่เกาะกันของผนังเซลล์นั้นอาจจะถูกทำลายในสภาพที่ pH ต่ำ หรืออาจจะเป็น pH ที่เหมาะสมสำหรับเอนไซม์ ที่จะทำให้ผนังเซลล์เปลี่ยนไป

86
การตอบสนองของพืชต่อออกซิน
1. การตอบสนองในระดับเซลล์ออกซินทำให้เกิด การขยายตัวของเซลล์ (Cell enlargement) 2. การตอบสนองของอวัยวะหรือพืชทั้งต้น 2.1 เกี่ยวข้องกับการตอบสนองของพืชต่อแสง (Phototropism) Geotropism ดังได้กล่าวมาแล้ว 2.2 การที่ตายอดข่มไม่ให้ตาข้างเจริญเติบโต (Apical Dominance)
 2. การตอบสนองของอวัยวะหรือพืชทั้งต้น. 2.1 เกี่ยวข้องกับการตอบสนองของพืชต่อแสง (Phototropism) Geotropism ดังได้กล่าวมาแล้ว. 2.2 การที่ตายอดข่มไม่ให้ตาข้างเจริญเติบโต (Apical Dominance)")
87
2.3 การติดผล เช่น กรณีของมะเขือเทศ ออกซินในรูปของ 4 CPA จะเร่งให้เกิดผลแบบ Pathenocarpic และในเงาะถ้าใช้ NAA 4.5 เปอร์เซ็นต์ จะเร่งการเจริญของเกสรตัวผู้ทำให้สามารถผสมกับเกสรตัวเมียได้ ในดอกที่ได้รับ NAA เกสรตัวเมียจะไม่เจริญเพราะได้รับ NAA ที่มีความเข้มข้นสูงเกินไป แต่เกสรตัวผู้ยังเจริญได้ ทำให้การติดผลเกิดมากขึ้น 2.4 ป้องกันการร่วงของผลโดยออกซินจะยับยั้งไม่ให้เกิด Abcission layer ขึ้นมา เช่น การใช้ 2,4-D ป้องกันผลส้มไม่ให้ร่วง หรือ NAA สามารถป้องกันการร่วงของผลมะม่วง 2.5 ป้องกันการร่วงของใบ 2.6 ในบางกรณีออกซินสามารถทำให้สัดส่วนของดอกตัวเมีย และตัวผู้เปลี่ยนไปโดยออกซินจะกระตุ้นให้มีดอกตัวเมียมากขึ้น

88
จิบเบอเรลลิน (Gibberellins)
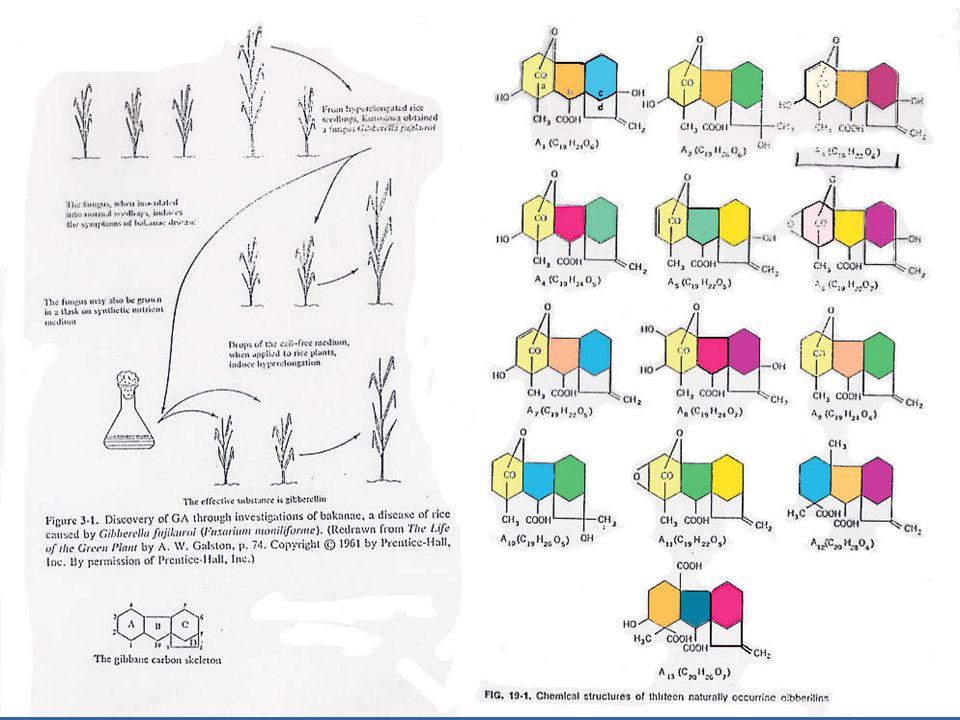
เมื่อ Kurosawa นักวิทยาศาสตร์ชาวญี่ปุ่น ศึกษาในต้นข้าวที่เป็นโรค Bakanae หรือโรคข้าวตัวผู้ ซึ่งเกิดจากเชื้อรา Gibberella fujikuroi หรือ Fusarium moniliforme ซึ่งทำให้ต้นข้าวมีลักษณะสูงกว่าต้นข้าวปกติ ทำให้ล้มง่าย ความรู้เกี่ยวกับโครงสร้างและส่วนประกอบทางเคมีของจิบเบอเรลลินนั้นได้รับการศึกษาในปี 1954 โดยนักเคมีชาวอังกฤษซึ่งสามารถแยกสารบริสุทธิ์จากอาหารเลี้ยงเชื้อรา Gibberella fujikuroi และเรียกสารนี้ว่ากรดจิบเบอเรลลิค (Gibberellic Acid)
")
90
ในปัจจุบันมีจิบเบอเรลลินซึ่งเป็นชื่อเรียกทั่ว ๆ ไปของสารประกอบประเภทนี้ประมาณไม่น้อยกว่า 80 ชนิด ชื่อเรียกสารประกอบชนิดนี้จะตั้งชื่อดังนี้ คือ Gibberellins A1 (GA1), A2, A3 เป็นต้น โดยที่กรดจิบเบอเรลลิค คือ GA3 GA ทุกชนิดจะมีโครงสร้างพื้นฐานของโมเลกุลเป็น Gibberellane Carbon Skeleton ซึ่งจะเหมือนกับกรดจิบเบอเรลลิค จะแตกต่างกันตรงจำนวนและตำแหน่งของกลุ่มที่เข้าแทนที่ในวงแหวนและระดับของความอิ่มตัวของวงแหวน A GA ประกอบด้วยคาร์บอนประมาณ 19-20 อะตอม ซึ่งจะรวมกันเป็นวงแหวน 4 หรือ 5 วงและจะต้องมีกลุ่มคาร์บอก-ซิล อย่างน้อย 1 กลุ่ม โดยใช้ชื่อย่อว่า GA

91
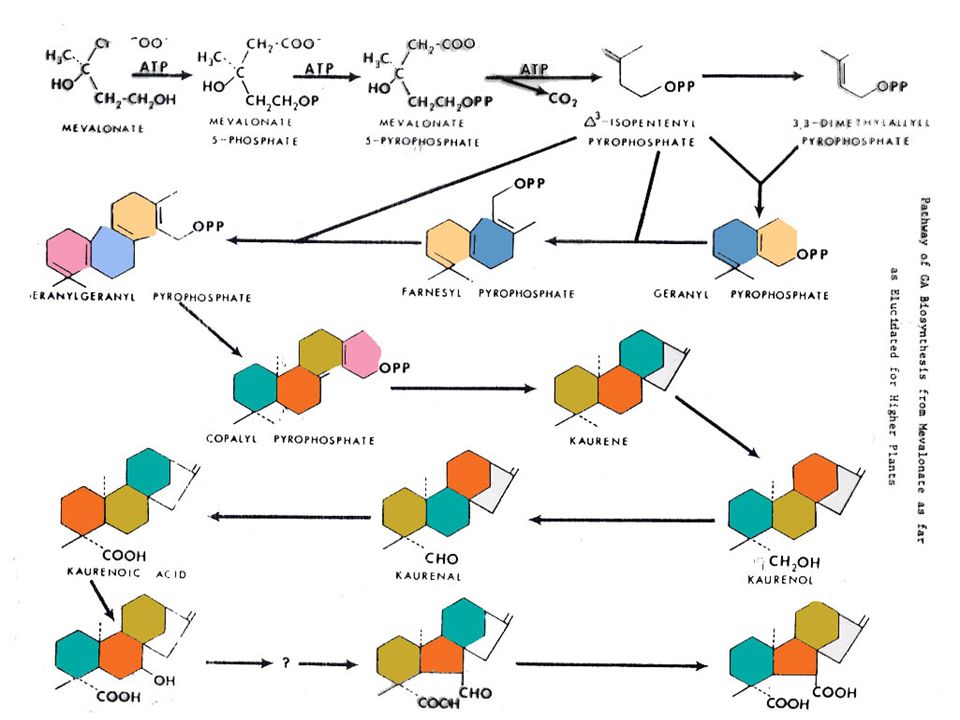
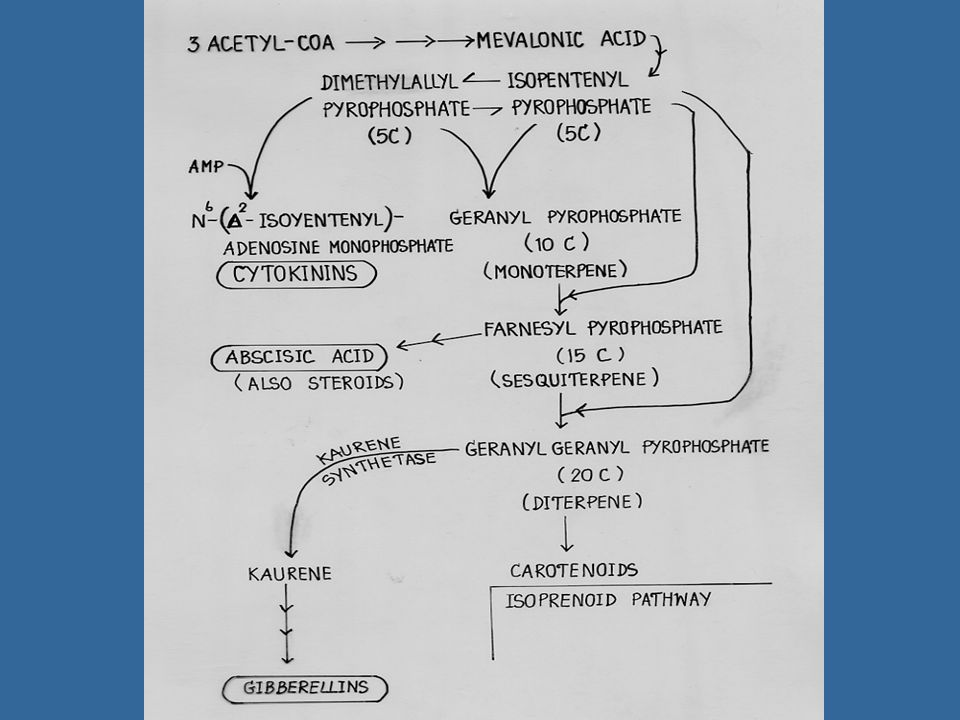
การสังเคราะห์จิบเบอเรลลิน
การสังเคราะห์ GA แสดงอยู่ในรูปที่ โดยที่สารเริ่มต้นเป็นกรดเมวาโลนิค (Mevalonic Acid) เปลี่ยนไปตามวิถีจนเกิดเป็นกรดคอรีโนอิค (Kaurenoic Acid) แล้วจึงเปลี่ยนไปเป็น GA ซึ่งวิถีในช่วงที่เปลี่ยนไปเป็น GA ชนิดต่าง ๆ นี้ยังไม่ทราบแน่ชัดนัก สารชนิดแรกที่มีวงแหวนของ Gibberellane คือ อัลดีไฮด์ของ GA12
 เปลี่ยนไปตามวิถีจนเกิดเป็นกรดคอรีโนอิค (Kaurenoic Acid) แล้วจึงเปลี่ยนไปเป็น GA ซึ่งวิถีในช่วงที่เปลี่ยนไปเป็น GA ชนิดต่าง ๆ นี้ยังไม่ทราบแน่ชัดนัก สารชนิดแรกที่มีวงแหวนของ Gibberellane คือ อัลดีไฮด์ของ GA12.")
92
การสังเคราะห์จิบเบอเรลลิน
ในปัจจุบันมีสารชะงักการเจริญเติบโต เช่น CCC หรือ Cycocel AM Phosfon-D และ SADH หรือ Alar ซึ่งใช้กันมากในการเกษตร สารเหล่านี้บางชนิดสามารถระงับ กระบวนการสังเคราะห์จิบเบอเรลลินได้ เช่น AMO-1618 สามารถระงับการสังเคราะห์จิบเบอเรลลินในอาหารสำรอง (Endosperm) ของแตงกวาป่า โดยระงับในช่วงการเปลี่ยน Geranylgeranyl pyrophosphate ไปเป็น Kaurene ในทำนองเดียวกัน CCC สามารถระงับกระบวนการนี้ได้ด้วย
 ของแตงกวาป่า โดยระงับในช่วงการเปลี่ยน Geranylgeranyl pyrophosphate ไปเป็น Kaurene ในทำนองเดียวกัน CCC สามารถระงับกระบวนการนี้ได้ด้วย.")
97
การสังเคราะห์จิบเบอเรลลิน
ใบอ่อน ผลอ่อนและต้นอ่อนเป็นส่วนที่สร้าง GA ของพืช รากพืชอาจจะสามารถสร้าง GA ได้บ้าง GA ที่พบในปัจจุบันจึงเป็นสารธรรมชาติทั้งสิ้น

98
การสลายตัวของจิบเบอเรลลิน
จิบเบอเรลลินมีกิจกรรมทางสรีรวิทยาอยู่ได้เป็นระยะเวลานานในเนื้อเยื่อพืช จิบเบอเรลลินสามารถเปลี่ยนจากชนิดหนึ่งไปเป็นจิบเบอเรลลินอีกชนิดหนึ่งได้ในเนื้อเยื่อพืช จิบเบอเรลลินในรูปของไกลโคไซด์ (Glycosides) กรดจิบเบอเรลลิค ซึ่งอยู่ในสภาพสารละลายถูกทำให้สลายตัวได้โดยใช้ Acid Hydrolysis ที่อุณหภูมิสูงและได้ผลิตภัณฑ์คือกรดจิบเบอเรลลี-นิค (Gibberellenic Acid) และกรดจิบเบอริค (Gibberic Acid)
 กรดจิบเบอเรลลิค ซึ่งอยู่ในสภาพสารละลายถูกทำให้สลายตัวได้โดยใช้ Acid Hydrolysis ที่อุณหภูมิสูงและได้ผลิตภัณฑ์คือกรดจิบเบอเรลลี-นิค (Gibberellenic Acid) และกรดจิบเบอริค (Gibberic Acid)")
99
การหาปริมาณจิบเบอเรลลิน
1. ใช้วิธีโครมาโตกราฟ เช่น GC หรือ Gas Chromatograph และ Paper Chromatograph 2. ใช้วิธี Bioassay โดยการที่จิบเบอเรลลินสามารถทำให้พืชแคระ (ข้าวโพดและถั่ว) เจริญเป็นต้นปกติได้ หรือโดยการที่จิบเบอเรลลินสามารถป้องกันการเกิดการเสื่อมสลาย (Senescence) หรือโดยหาปริมาณจิบเบอเรลลินจากการกระตุ้นให้เมล็ดข้าว-บาร์เลย์สร้างเอนไซม์ แอลฟา อะมัยเลส ( -amylase) ในอาหารสำรอง
 เจริญเป็นต้นปกติได้ หรือโดยการที่จิบเบอเรลลินสามารถป้องกันการเกิดการเสื่อมสลาย (Senescence) หรือโดยหาปริมาณจิบเบอเรลลินจากการกระตุ้นให้เมล็ดข้าว-บาร์เลย์สร้างเอนไซม์ แอลฟา อะมัยเลส ( -amylase) ในอาหารสำรอง.")
100
จิบเบอเรลลินสามารถเคลื่อนย้ายหรือเคลื่อนที่ในพืชได้ทั้งทางเบสิพีตัล และอะโครพีตัล และการเคลื่อนที่ไม่มีโพลาริตี้ การเคลื่อนย้ายเกิดขึ้นทั้งในส่วนของท่ออาหารและท่อน้ำ จิบเบอเรลลินจากยอดอ่อนลงมาสู่ส่วนล่างของลำต้นนั้นไม่ได้เกิดในท่อน้ำ ท่ออาหารเพราะส่วนของยอดอ่อนเป็นส่วนที่ดึงอาหารและธาตุอาหารให้เคลื่อนที่ขึ้นไปแบบอะโครพีตัล ดังนั้นจิบเบอเรลลิน จึงไม่ได้เคลื่อนที่ทางท่ออาหารและยังไม่ทราบวิถีการเคลื่อนที่แน่ชัด

101
กลไกในการทำงานของจิบเบอเรลลิน
ระดับของกิจกรรมของเอนไซม์หลายชนิดมีผลกระทบจากปริมาณของจิบเบอเรลลิน เอนไซม์ซึ่งมีกิจกรรมเพิ่มขึ้นเมื่อได้รับจิบเบอเรลลิน คือ เอนไซม์ แอลฟาและเบตา-อะมัยเลส ( และ -amylase) โปรตีเอส (Protease) และไรโบนิวคลีเอส (Ribonuclease) ซึ่งพบในเมล็ดข้าวบาร์เลย์ซึ่งกำลังงอก
 โปรตีเอส (Protease) และไรโบนิวคลีเอส (Ribonuclease) ซึ่งพบในเมล็ดข้าวบาร์เลย์ซึ่งกำลังงอก.")
102
ระดับกิจกรรมของเอนไซม์ซึ่งถูกควบคุมโดยจิบเบอ-เรลลิน ทำกันมากในเอนไซม์ แอลฟา อะมัยเลส ในเมล็ดข้าวบาร์เลย์ ในเมล็ดข้าวบาร์เลย์ที่แห้งที่ยังไม่ดูดซับน้ำจะไม่มีเอนไซม์แอลฟา อะมัยเลส ปรากฏอยู่เอนไซม์นี้จะปรากฏขึ้นและปลดปล่อยออกมาจากชั้นของอะลีโรนของเมล็ด เป็นการตอบสนองต่อจิบเบอ-เรลลินซึ่งสังเคราะห์จากต้นอ่อนที่กำลังงอก เนื้อเยื่อชั้นอะลีโรนซึ่งแยกจากเมล็ดที่ไม่งอกจะมีกิจกรรมของแอลฟา อะมัยเลส น้อยมาก

103
จิบเบอเรลลินควบคุมกิจกรรมของแอลฟา อะมัยเลส ผ่านทางการสังเคราะห์ RNA สารชะงักการสังเคราะห์ RNA เช่น แอคติโนมัยซิน-ดี (Actinomycin-D) จะชะงักกระบวนการกระตุ้นการสังเคราะห์ RNA 2-3 ชั่วโมง หลังจากเติมจิบเบอเรลลิน ในขณะที่สารชะงักการสังเคราะห์โปรตีน เช่น ไซโคลเฮคซิไมด์ (Cycloheximide) จะระงับการปรากฏของกิจกรรมของแอลฟา อะมัยเลส หลังจากช่วง "lag" เริ่มต้น
 จะชะงักกระบวนการกระตุ้นการสังเคราะห์ RNA 2-3 ชั่วโมง หลังจากเติมจิบเบอเรลลิน ในขณะที่สารชะงักการสังเคราะห์โปรตีน เช่น ไซโคลเฮคซิไมด์ (Cycloheximide) จะระงับการปรากฏของกิจกรรมของแอลฟา อะมัยเลส หลังจากช่วง lag เริ่มต้น")
104
บทบาทของจิบเบอเรลลินที่มีต่อพืช
1. กระตุ้นการเจริญเติบโตของพืชทั้งต้น 2. กระตุ้นการงอกของเมล็ดที่พักตัวและตาที่พักตัว 3. การแทงช่อดอก การออกดอกของพืชเกี่ยวข้องกับปัจจัยหลายอย่าง เช่น อายุ และสภาพแวดล้อม จิบเบอเรลลินสามารถแทนความต้องการวันยาวในพืชบางชนิดได้ และยังสามารถทดแทนความต้องการอุณหภูมิต่ำ (Vernalization) ในพืชพวกกะหล่ำปลี และแครอท
 ในพืชพวกกะหล่ำปลี และแครอท.")
105
4. จิบเบอเรลลิน สามารถกระตุ้นการเคลื่อนที่ของอาหารในเซลล์สะสมอาหารหลังจากที่เมล็ดงอกแล้ว
5. กระตุ้นให้เกิดผลแบบ Parthenocarpic ในพืชบางชนิด เปลี่ยนรูปร่างของใบพืชบางชนิด เช่น English Ivy และทำให้พืชพัฒนาการเพื่อทนความเย็นได้ 6. จิบเบอเรลลินมักเร่งให้เกิดดอกตัวผู้

107
ไซโตไคนิน (Cytokinins)
1920 Haberlandt ได้แสดงให้เห็นว่ามีสารชนิดหนึ่งเกิดอยู่ในเนื้อเยื่อพืชและกระตุ้นให้เนื้อเยื่อพาเรนไคมาในหัวมันฝรั่งกลับกลายเป็นเนื้อเยื่อเจริญได้ ซึ่งแสดงว่าสารชนิดนี้สามารถกระตุ้นให้มีการแบ่งเซลล์ ต่อมามีการพบว่าน้ำมะพร้าวและเนื้อเยื่อของหัวแครอทมีคุณสมบัติในการกระตุ้นการแบ่งเซลล์เช่นกัน

108
ไซโตไคนิน (Cytokinins)
Skoog และ Steward ทำการทดลองในสหรัฐอเมริกา โดยศึกษาความต้องการสิ่งที่ใช้ในการเจริญเติบโตของกลุ่มก้อนของเซลล์ (Callus) ซึ่งเป็นเซลล์ที่แบ่งตัวอย่างรวดเร็ว แต่ไม่มีการเปลี่ยนแปลงทางคุณภาพเกิดขึ้นของ pith จากยาสูบและรากของแครอท
 ซึ่งเป็นเซลล์ที่แบ่งตัวอย่างรวดเร็ว แต่ไม่มีการเปลี่ยนแปลงทางคุณภาพเกิดขึ้นของ pith จากยาสูบและรากของแครอท.")
109
ไซโตไคนิน (Cytokinins)
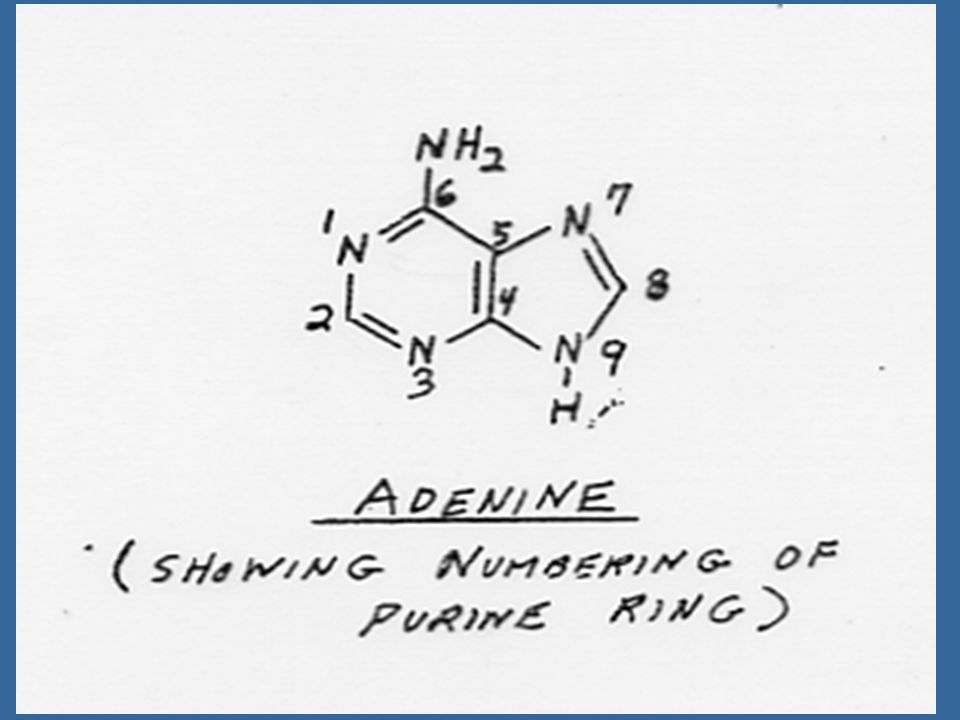
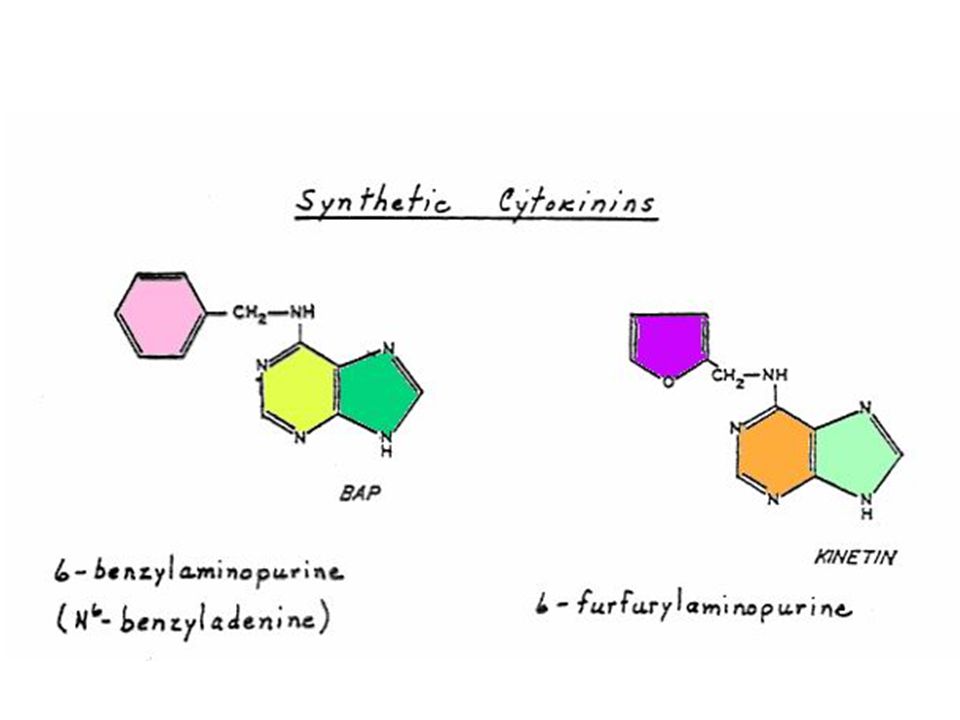
Miller ได้แยกสารอีกชนิดหนึ่งซึ่งมีคุณสมบัติคล้ายคลึงแต่มีประสิทธิภาพดีกว่าอะดีนีน ซึ่งได้จากการสลายตัวของ DNA ของสเปิร์มจากปลาแฮร์ริง สารชนิดนี้ คือ 6-(furfuryl-amino) purine ซึ่งมีสูตรโครงสร้างคล้ายอะดีนีน เนื่องจากสารชนิดนี้สามารถกระตุ้นให้เกิดการแบ่งเซลล์โดยร่วมกับ ออกซิน จึงได้รับชื่อว่าไคเนติน (Kinetin)
 purine ซึ่งมีสูตรโครงสร้างคล้ายอะดีนีน เนื่องจากสารชนิดนี้สามารถกระตุ้นให้เกิดการแบ่งเซลล์โดยร่วมกับ ออกซิน จึงได้รับชื่อว่าไคเนติน (Kinetin)")
110
สารสังเคราะห์ที่มีกิจกรรมของไซโตไคนินสูงที่สุดคือ เบนซิลอะดีนีน (Benzyladenine หรือ BA) และเตตระไฮโดรไพรานีลเบนซิลอะดีนีน (tetrahydropyranylbenzyladenine หรือ PBA)
 และเตตระไฮโดรไพรานีลเบนซิลอะดีนีน (tetrahydropyranylbenzyladenine หรือ PBA)")
111
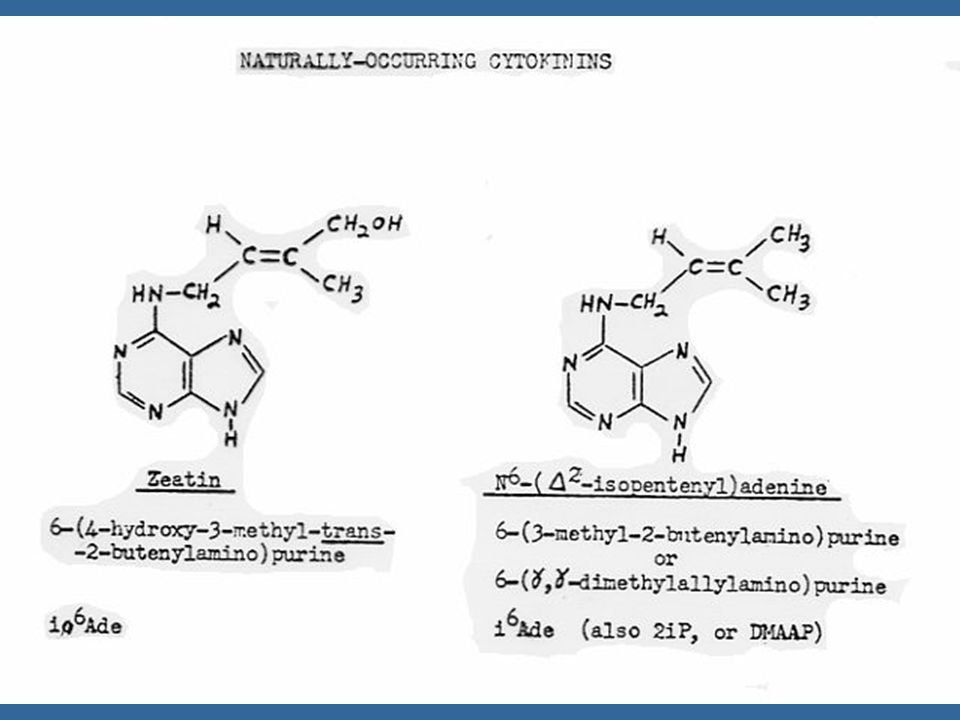
ไซโตไคนินที่เกิดในธรรมชาติเป็นสารประกอบพิวรีน ในปี Letham ได้แยกไซโตไคนินชนิดหนึ่งจากเมล็ดข้าวโพดหวาน และพบว่า เป็นสาร 6-(4-hydroxy-3-methyl but-2-enyl) aminopurine ซึ่ง Letham ได้ตั้งชื่อว่า ซีเอติน (Zeatin)
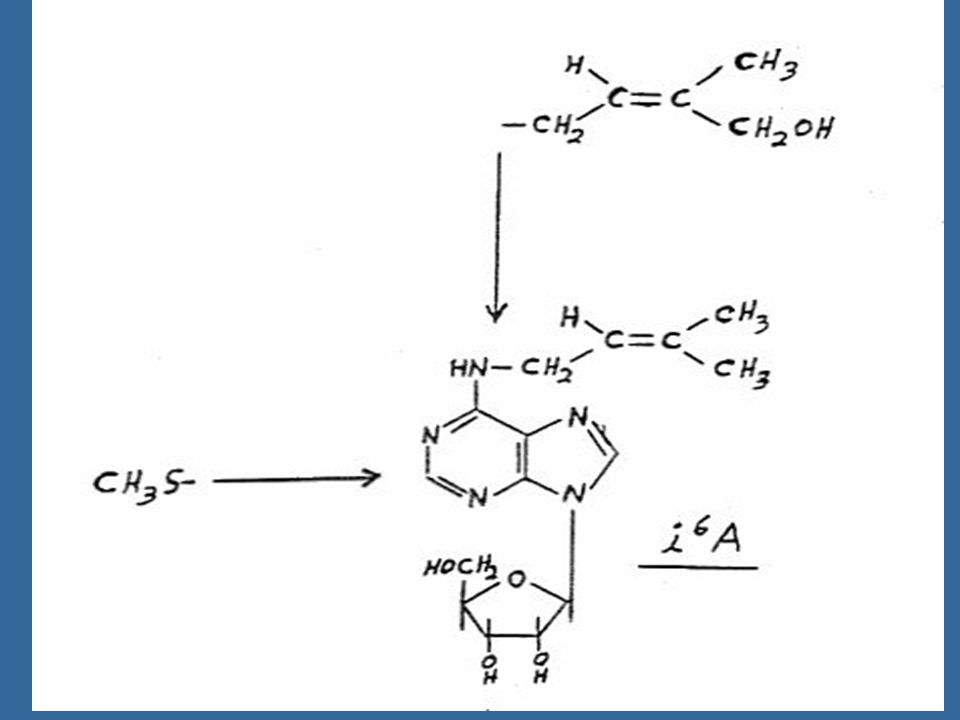
 aminopurine ซึ่ง Letham ได้ตั้งชื่อว่า ซีเอติน (Zeatin)")
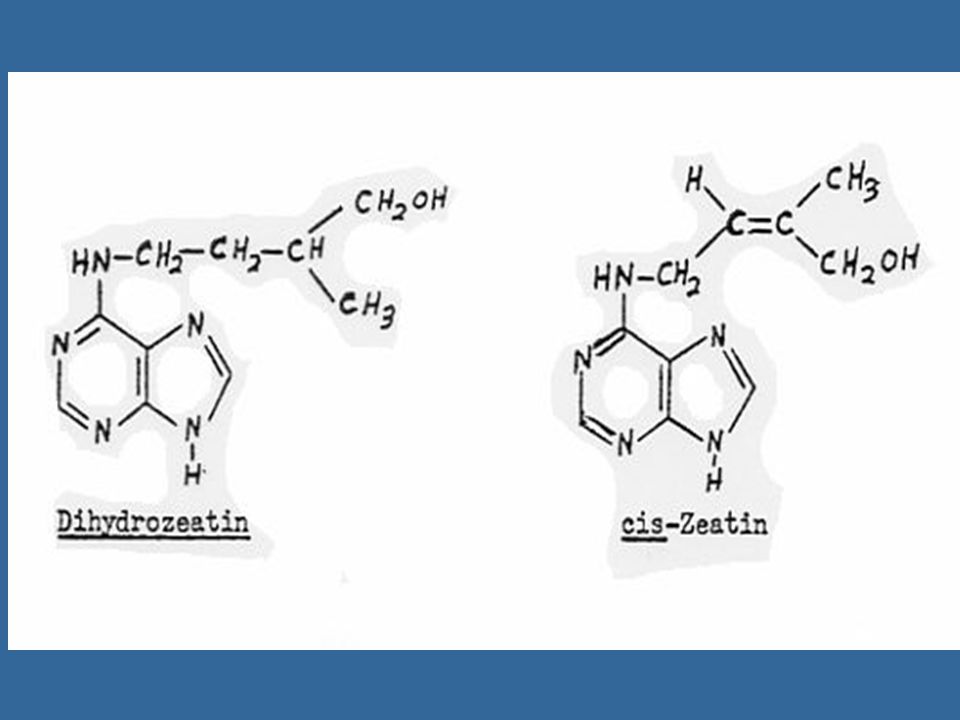
112
นับตั้งแต่มีการแยกไซโตไคนินชนิดแรกคือซีเอตินแล้ว ก็มีการค้นพบไซโตไคนิน อีกหลายชนิดซึ่งทุกชนิดเป็นอนุพันธ์ของอะดีนีน คือ เป็น 6-substituted amino purines ซีเอตินเป็นไซโต-ไคนินธรรมชาติซึ่งมีประสิทธิภาพสูงที่สุด

117
Non-purine Compounds with Cytokinin-like Activity

123
The End