ดาวน์โหลดงานนำเสนอ
งานนำเสนอกำลังจะดาวน์โหลด โปรดรอ

1
Posttranslational Modifications
Protein Synthesis : Translation and Posttranslational Modifications รัชนีกร กัลล์ประวิทธ์ 2557

2
Protein synthesis Translation
คล้ายคลึงกันทั้งใน prokaryote และ eukaryote สารตั้งต้น คือ กรดอะมิโน 20 ชนิด ชนิดของโปรตีนที่สังเคราะห์ ถูกกำหนดโดย mRNA การสังเคราะห์โปรตีนเริ่มจากจุดเริ่มต้นทางด้าน NH2-terminal ไปสิ้นสุดที่ COOH-terminal ในมนุษย์สามารถพบการสังเคราะห์โปรตีนได้ทั้งใน cytosol, RER และ mitochondria

3
ปัจจัยที่ใช้ในการสังเคราะห์โปรตีน
L-amino acid t-RNA Genetic code Aminoacyl-tRNA synthetase ribosome

4
Amino acid

5
Transfer RNA (tRNA) มีประมาณ 50 ชนิด โดยกรดอะมิโนบางชนิดมี tRNA ที่เป็นคู่ มากกว่า 1 ชนิด Clover leaf หรือ L-shape tRNA จะมีเบส 3 ตัวที่ complementary กับ codon บน mRNA เรียกเบส 3 ตัวนี้ว่า anticodon Wobble position

6
Transfer RNA (tRNA)
")
7
Wobble position Wobble position (base) คือ เบสที่อยู่ตำแหน่ง 3’ ของ codon และเบสตำแหน่ง 5’ ของ anticodon ซึ่งโดยทั่วไป การจับคู่กัน ของเบสคู่นี้อาจไม่เป็นไปตามกฎทั่วไปของการจับคู่ การมี wobble base ทำให้ tRNA 1 ชนิด จับกับ codon ของ mRNA ได้มากกว่า 1 codon 5’ base of anticodon 3’ base of codon C A U G I A or G C or U U or C or A
 คือ เบสที่อยู่ตำแหน่ง 3’ ของ codon และเบสตำแหน่ง 5’ ของ anticodon ซึ่งโดยทั่วไป การจับคู่กัน ของเบสคู่นี้อาจไม่เป็นไปตามกฎทั่วไปของการจับคู่ การมี wobble base ทำให้ tRNA 1 ชนิด จับกับ codon ของ mRNA ได้มากกว่า 1 codon. 5’ base of anticodon. 3’ base of codon. C. A. U. G. I. A or G. C or U. U or C or A.")
8
Genetic code U C A G UUU UAU UGU UCU Tyr Cys UUC Phe UGC U C A G UCC
Second position U C A G UUU UAU UGU UCU Tyr Cys UUC Phe UGC U C A G UCC UAC U Ser UUA UCA UAA UGA STOP Leu STOP UUG UCG UAG UGG Trp CUU First position His CCU CCC CCA CCG CAU CAC CGU CGC CGA CGG U C A G CUC Leu C Pro Arg CUA Third position Gln CUG CAG CAA AUU AUC AUA Asn Ser lle ACU ACC ACA ACG AGU AGC U C A G AAC AAU Thr A Lys AUG Met Arg AAG AAA AGG AGA GUU GUA GUG Asp GCU GCC GCA GCG GAU GAC U C A G GUC GGU GGC GGA GGG Gly G Val Ala Glu GAG GAA

9
Genetic code รหัสพันธุกรรม
แต่ละรหัสประกอบด้วย nucleotide 3 ตัว เรียงต่อกัน เรียกว่า triplet code หรือ codon สิ่งมีชีวิตทุกชนิด ทั้ง prokaryote และ eukaryote ใช้ genetic code เดียวกัน (ยกเว้นใน mitochondria ของมนุษย์มีรหัสบางรหัส ต่างออกไป) จึงอาจเรียกว่า universal genetic code Genetic code บน mRNA จะมีแต่ละ codon เรียงต่อกันไป โดยไม่มี การเว้นวรรค (commaless) และไม่ซ้อนกัน (non-overlapping)
 จึงอาจเรียกว่า universal genetic code. Genetic code บน mRNA จะมีแต่ละ codon เรียงต่อกันไป โดยไม่มี การเว้นวรรค (commaless) และไม่ซ้อนกัน (non-overlapping)")
10
Genetic code รหัสพันธุกรรมมีทั้งหมด 64 รหัส (43 = 64) โดย 61 รหัสจะเป็นรหัส สำหรับกรดอะมิโน 20 ชนิด ที่เหลือ 3 รหัส เป็นรหัสสำหรับหยุดการ สังเคราะห์โปรตีน กรดอะมิโนส่วนใหญ่จะมีรหัสพันธุกรรมมากกว่า 1 รหัส เรียก degenerate มีกรดอะมิโน 2 ชนิด (methionine และ tryptophan) ที่มีรหัสพันธุกรรม รหัสเดียว AUG เป็นรหัสพันธุกรรมสำหรับ methionine และเป็นรหัสเริ่มต้นการ สังเคราะห์โปรตีน เรียกว่า initiation codon UAA, UAG และ UGA เป็นรหัสที่หยุดการสังเคราะห์โปรตีน เรียกว่า nonsense (stop) codon
 โดย 61 รหัสจะเป็นรหัส สำหรับกรดอะมิโน 20 ชนิด ที่เหลือ 3 รหัส เป็นรหัสสำหรับหยุดการ สังเคราะห์โปรตีน. กรดอะมิโนส่วนใหญ่จะมีรหัสพันธุกรรมมากกว่า 1 รหัส เรียก degenerate. มีกรดอะมิโน 2 ชนิด (methionine และ tryptophan) ที่มีรหัสพันธุกรรม รหัสเดียว. AUG เป็นรหัสพันธุกรรมสำหรับ methionine และเป็นรหัสเริ่มต้นการ สังเคราะห์โปรตีน เรียกว่า initiation codon. UAA, UAG และ UGA เป็นรหัสที่หยุดการสังเคราะห์โปรตีน เรียกว่า nonsense (stop) codon.")
11
Gene mutation & protein synthesis
Regulatory element mutation : ทำให้การเกิด transcription เปลี่ยนแปลง (เพิ่มขึ้น/ลดลง) ส่งผลให้ปริมาณของโปรตีนเปลี่ยนไป Structural gene mutation : ทำให้เกิดการเปลี่ยนแปลงต่าง ๆ ดังนี้ Missense mutation Nonsense mutation Chain termination (sense) mutation Splicing mutation Silent mutation Frameshift mutation
 ส่งผลให้ปริมาณของโปรตีนเปลี่ยนไป. Structural gene mutation : ทำให้เกิดการเปลี่ยนแปลงต่าง ๆ ดังนี้ Missense mutation. Nonsense mutation. Chain termination (sense) mutation. Splicing mutation. Silent mutation. Frameshift mutation.")
12
Structural gene mutation
Missense mutation : โปรตีนมีกรดอะมิโนเปลี่ยนไป CAU (His) CAG (Gln) Nonsense mutation : โปรตีนสั้นลง (truncated polypeptide chain) CGA (Arg) UGA (stop) Sense mutation : โปรตีนยาวขึ้น UAA (stop) UAC (Tyr)
 CAG (Gln) Nonsense mutation : โปรตีนสั้นลง (truncated polypeptide chain) CGA (Arg) UGA (stop) Sense mutation : โปรตีนยาวขึ้น. UAA (stop) UAC (Tyr)")
13
Structural gene mutation
Silent mutation AAA (Lys) AAG (Lys) Frameshift mutation Pro Gln Trp Ser Arg His Normal 5' 3′ C C A C A G U G G A G U C G A C A U U Mutant 5' 3′ C C A C A G U G G A A G U C G A C A U (base insertion ) Pro Gln Trp Lys Ser Thr
 AAG (Lys) Frameshift mutation. Pro. Gln. Trp. Ser. Arg. His. Normal 5 3′ C C A C A G U G G A G U C G A C A U U. Mutant 5 3′ C C A C A G U G G A A G U C G A C A U. (base insertion ) Pro. Gln. Trp. Lys. Ser. Thr.")
14
Aminoacyl-tRNA synthetase
ทำหน้าที่จับคู่กรดอะมิโนกับ tRNA ที่เป็นคู่กัน (cognate tRNA) เรียกขั้นตอนนี้ว่า activation of amino acid amino acid + ATP + tRNA aminoacyl-tRNA + AMP + PPi PPi Pi ขบวนการนี้อาศัย Mg2+ เป็น cofactor และต้องการ ATP (2 ATP) เอนไซม์นี้มี 20 ชนิด แต่ละชนิดจำเพาะกับกรดอะมิโนแต่ละตัว
 เรียกขั้นตอนนี้ว่า activation of amino acid. amino acid + ATP + tRNA aminoacyl-tRNA + AMP + PPi. PPi 2Pi. ขบวนการนี้อาศัย Mg2+ เป็น cofactor และต้องการ ATP (2 ATP) เอนไซม์นี้มี 20 ชนิด แต่ละชนิดจำเพาะกับกรดอะมิโนแต่ละตัว.")
15
Aminoacyl-tRNA synthetase
การจับกันของกรดอะมิโนกับ tRNA เกิดจาก α-COOH group ของ กรดอะมิโนกับ 3’-OH group ของ tRNA จับกัน เกิดเป็น high energy ester bond ซึ่งพลังงานของพันธะนี้จะใช้ในการสังเคราะห์ peptide bond ต่อไป เอนไซม์นี้ยังสามารถ proofreading ตรวจสอบ aa-tRNA ที่เกิดขึ้น โดยถ้า aa-tRNA มีโครงสร้างที่ผิด เอนไซม์จะสลาย aa-tRNA กลับเป็นกรดอะมิโนและ tRNA

16
Ribosome ตำแหน่งที่เกิดการสังเคราะห์โปรตีน
ประกอบด้วย 2 subunits (small & large subunit) large subunit ประกอบด้วย P site : เป็นบริเวณที่เกิดสาย polypeptide A site : เป็นบริเวณที่ aa-tRNA มาเกาะ Prokaryotic ribosome : 70S (50S & 30S) Eukaryotic ribosome : 80S (60S & 40S)
 large subunit ประกอบด้วย. P site : เป็นบริเวณที่เกิดสาย polypeptide. A site : เป็นบริเวณที่ aa-tRNA มาเกาะ. Prokaryotic ribosome : 70S (50S & 30S) Eukaryotic ribosome : 80S (60S & 40S)")
17
Process of translation
Initiation Elongation Termination

18
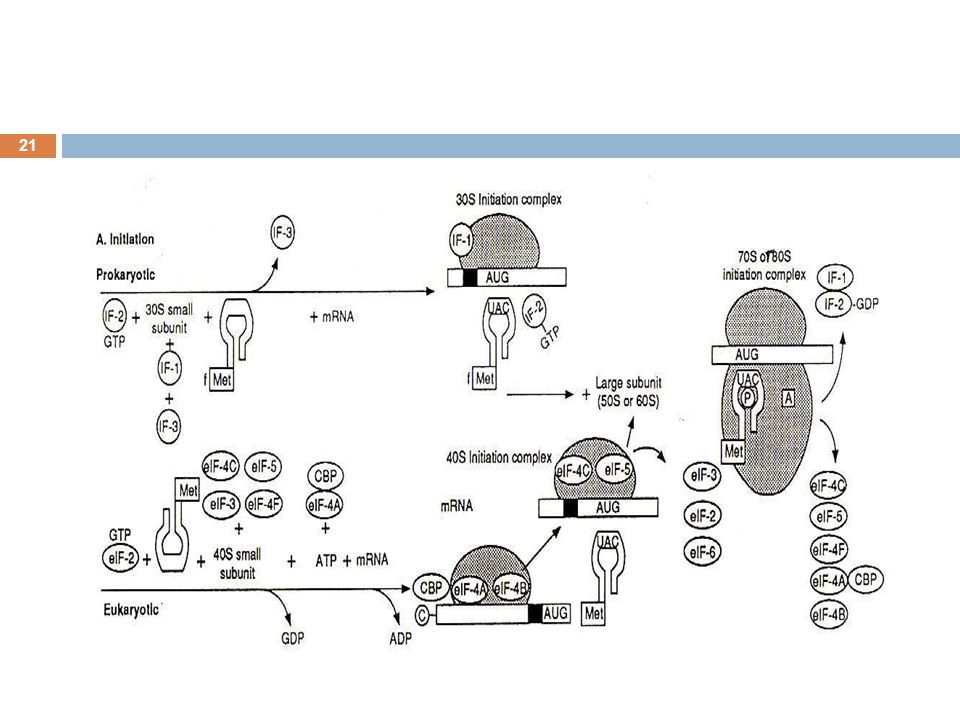
Initiation การ identify จุดเริ่มต้นของ coding sequence บน mRNA (ซึ่งก็คือ codon AUG ตัวแรกจากด้าน 5’) ปัจจัยที่ใช้ในขั้นตอน initiation mRNA Met-tRNA (ใน prokaryote ใช้ formyl methionyl-tRNA) ribosome initiation factors (IF) GTP ATP
 ribosome. initiation factors (IF) GTP. ATP.")
19
Process of initiation (eukaryotes)
eIF2a จับกับ GTP และ Met-tRNAi ได้เป็น ternary complex Ribosome แยกเป็น 2 subunit โดย 40S subunit รวมกับ eIF3, ternary complex และ eIF4c ได้เป็น eIF2a.Met- tRNAi.GTP, eIF3.40S, eIF4c complex complex นี้จะจับ mRNA ทางด้าน 5’ ได้เป็น preinitiation complex ซึ่งต้องอาศัย ATP และ IF ในการอ่านรหัสจากด้าน 5’ ไปด้าน 3’ จนถึง codon AUG ชุดแรก

20
Process of initiation (eukaryotes)
anticodon ของ tRNA เกิด base pairing กับ AUG ของ mRNA ทำให้ GTP ถูกเปลี่ยนเป็น GDP IF ต่าง ๆ และ GDP ถูกปล่อยออกมา และ 60S subunit ของ ribosome เข้ามารวม เกิดเป็น initiation complex

22
Process of initiation (prokaryotes)
มีลำดับของพิวรีนนิวคลีโอไทด์ที่เรียกว่า Shine-Dalgarno sequence อยู่ประมาณ 10 นิวคลีโอไทด์ ทางด้าน upstream ของ AUG IF ต่าง ๆ น้อยกว่า ใน eukaryote AUG 5' 3' mRNA Prok. Shine-Dalgarno sequence ( 10 nt. upstream ของ AUG ) initiation complex อยู่ที่ P site ของ ribosome
 initiation complex อยู่ที่ P site ของ ribosome.")
23
Elongation เกิดขึ้นหลังการเกิด initiation complex
ribosome จะต่อสาย peptide ให้ยาวขึ้นโดยการอ่าน codon บน mRNA ไปเรื่อย ๆ จนถึง termination codon ปัจจัยที่ใช้ คือ initiation complex aminoacyl-tRNA ที่รวมกับ eEF1α และ GTP เป็น complex elongation factors (EF) GTP
 GTP.")
24
Process of elongation eEF-1α ที่จับกับ GTP จะพา aa-tRNA ไปรวมกับ initiation complex ที่ A site เกิด base pairing ระหว่าง base ของ codon บน mRNA กับ anticodon บน aa-tRNA ทำให้ eEF-1α กับ GTP เปลี่ยนเป็น GDP. eEF-1α ต่อมา GDP หลุดออกจาก ribosome ปล่อย aa- tRNA ไว้ที่ A site ของ ribosome peptidyl transferase ใน ribosome สลาย ester bond ระหว่างกรดอะมิโนกับ tRNA ที่ P site แล้วนำ α-COOH group มาต่อกับ α-NH2 group ของกรดอะมิโนอีกตัวที่ A site

25
Process of elongation เกิด translocation ย้าย peptidyl-tRNA จาก A site มาที่ P site โดยอาศัย eEF-2 (translocase) และ GTP deacylated tRNA จะเคลื่อนที่จาก P site ไป E site เมื่อ A site ว่าง aa-tRNA ตัวใหม่จะเข้ามาจับ และเกิด ขบวนการต่าง ๆ ดังที่กล่าวมาใหม่

26
Process of elongation

27
Termination เกิดเมื่อ ribosome สังเคราะห์ peptide จนมาถึง termination codon (UAA, UAG หรือ UGA) eRF และ GTP จะจับที่ A site ปล่อย deacylated tRNA จาก E site ปล่อย peptide ที่สังเคราะห์ได้ออกจาก ribosome

28
Process of termination

29
Energy requirement for protein synthesis
Aminoacylation of tRNA ATP AMP + Ppi Binding of Met-tRNAi to P site Aminoacyl-tRNA binding to A site Translocation Termination

30
Polyribosome (polysome)
mRNA สายเดี่ยวแต่มี ribosome เกาะอยู่หลายกลุ่ม เกิดจากเมื่อ ribosome เริ่มสังเคราะห์โปรตีนไปแล้ว ribosome ตัวใหม่จะเริ่มสังเคราะห์โปรตีนอีกโมเลกุลได้โดยไม่ต้องรอให้ ribosome ตัวแรกสังเคราะห์โปรตีนจนจบสายก่อน การมี polyribosome ทำให้เซลล์สังเคราะห์โปรตีนได้เร็วขึ้น

31
Polyribosome

32
Inhibitors of protein synthesis
affect on prokaryotes : streptomycin, tetracycline, chloramphenicol & erythromycin affect on eukayotes : cycloheximide & diphtheria toxin affect both prokaryotes & eukaryotes : puromycin

33
Inhibitors of protein synthesis
Mechanism of action streptomycin binds to 30 s ribosomal subunit, preventing formation of the initiation complex tetracycline inhibits binding of aa-t RNA to A site chloramphenicol inhibits peptidyl transferase activity of 50s ribosomal subunit erythromycin binds to 50 s ribosomal subunit, prevents translocation cycloheximide inhibits peptidyl transferase in eukaryotes diphtheria toxin inhibit translocation of peptidyl-tRNA from A to P site puromycin prematurely terminates synthesis

34
Cycle repeated many times
Protein folding Hsp60 การสังเคราะห์โปรตีน ทำให้เกิด 1° structure ของโปรตีน สำหรับการเกิด 2° และ 3° structure โปรตีนบางชนิดสามารถเกิดได้เอง (self-assembly) แต่โปรตีนบางชนิด ต้องอาศัยโปรตีนชนิดอื่นเป็นตัวช่วย เช่น molecular chaperone (heat shock protein) ป้องกันการเกิด โครงสร้างที่ไม่ต้องการ และทำให้ โปรตีนเสถียร prolyl isomerase เปลี่ยน cis- trans isomer ของ proline ใน โปรตีน binding Cycle repeated many times
 แต่โปรตีนบางชนิด ต้องอาศัยโปรตีนชนิดอื่นเป็นตัวช่วย เช่น. molecular chaperone (heat shock protein) ป้องกันการเกิด โครงสร้างที่ไม่ต้องการ และทำให้ โปรตีนเสถียร. prolyl isomerase เปลี่ยน cis- trans isomer ของ proline ใน โปรตีน. binding. Cycle repeated many times.")
35
Protein folding mutation บางชนิด ทำให้การเกิด protein folding ผิดปกติไป และทำให้โปรตีนดังกล่าวไม่สามารถถูกส่งออกจาก ER ได้ ตัวอย่างเช่น mutation ของ α1-antiprotease (ทำหน้าที่เป็น elastase inhibitor) abnormal folding of α1-antiprotease retention of α1-antiprotease within hepatocyte elastase activity ในปอดสูงขึ้น lung destruction
 abnormal folding of α1-antiprotease. retention of α1-antiprotease within hepatocyte. elastase activity ในปอดสูงขึ้น. lung destruction.")
36
Protein translocation (protein targeting)
การเคลื่อนย้ายโปรตีนจากบริเวณที่สังเคราะห์ไปยังบริเวณที่ โปรตีนทำงาน แบ่งได้เป็น 2 กลุ่ม 1. Post-translational translocation 2. Cotranslational translocation

37
Post-translational translocation
การเคลื่อนย้ายโปรตีนที่เกิดหลังจากการสังเคราะห์โปรตีน เสร็จสิ้น เกิดที่ free ribosome พบในการสังเคราะห์โปรตีนเพื่อใช้ใน nucleus, cytosol, mitochondria และ peroxisome

38
Cotranslational translocation
การเคลื่อนย้ายโปรตีนที่เกิดขณะที่การสังเคราะห์โปรตีนยังไม่สิ้นสุด พบในการสังเคราะห์โปรตีนที่เป็นส่วนประกอบของ membrane และ secretory protein เช่น โปรตีนใน ER, Golgi body, lysosome โปรตีนในกลุ่มนี้จะมี signal sequence ที่ด้าน N-terminus ขั้นตอนการเกิด cotranslational translocation Signal sequence จะจับกับ signal recognition particle (SRP) SRP จะพาโปรตีนและ ribosome ไปจับกับ SRP receptor (docking protein) ที่ ER และโปรตีนจะถูกส่งเข้าไปใน lumen ของ ER เมื่อสังเคราะห์โปรตีนเสร็จ singal sequence จะถูกตัดออก โปรตีนใน lumen ถูกส่งไป Golgi body และเกิด exocytosis
 SRP จะพาโปรตีนและ ribosome ไปจับกับ SRP receptor (docking protein) ที่ ER และโปรตีนจะถูกส่งเข้าไปใน lumen ของ ER. เมื่อสังเคราะห์โปรตีนเสร็จ singal sequence จะถูกตัดออก. โปรตีนใน lumen ถูกส่งไป Golgi body และเกิด exocytosis.")
39
Cotranslational translocation

40
Secretion เป็นขบวนการส่งโปรตีนที่สังเคราะห์ ออกไปใช้นอกเซลล์ แบ่งได้เป็น 2 ประเภท Constitutive secretion ตัวอย่างเช่น การส่ง plasma protein ออกนอกเซลล์ Regulated excretion โปรตีนที่สังเคราะห์จะถูกเก็บไว้ใน secretory vesicle เมื่อเซลล์ถูกกระตุ้น โปรตีนจะถูกปล่อย ออกมาโดยวิธี exocytosis

41
Intracellular protein degradation
โดยทั่วไปโปรตีนแต่ละ ชนิดจะมี half-life ต่างกัน เช่น ในเซลล์ตับ เอนไซม์ ที่ควบคุม metabolism จะสลายเร็วกว่าโปรตีนอื่น โปรตีนที่มี PEST sequence (proline- glutamate-serine- threonine) มากจะมี half-life สั้น
 มากจะมี half-life สั้น.")
42
Intracellular protein degradation
การสลายโปรตีนในเซลล์ ATP dependent proteolysis เกิดขึ้นใน proteasomes โดยอาศัย โปรตีน ubiquitin Lysosome degradation Other proteolytic systems - Apoptosis หรือที่เรียกว่า programmed cell death อาศัยเอนไซม์ caspases - Calcium-dependent thiol proteases ที่เรียกว่า calpains

43
Ubiquitin-ATP dependent pathway
ทำหน้าที่สลายโปรตีนที่ผิดปกติ หรือหมดอายุการใช้งาน เช่น regulatory protein และ protein ที่เกี่ยวข้องกับการแบ่งตัว ของเซลล์ ubiquitin จะจับกับ NH3+ group ของ lysine ของโปรตีน ต้องการสลาย โดยอาศัย E1, E2 และ E3 จากนั้น protease complex จะสลายโปรตีนและปล่อย ubiquitin ออกไปใช้ได้ใหม่

44
Ubiquitin-ATP dependent protein degradation

45
Lysosome degradation เป็นการสลายโปรตีนโดย protease ของ lysosome ที่เรียกว่า cathepsins ซึ่งแบ่งออกตามหน้าที่ได้ดังนี้ Exocytosis เป็นการส่งเอนไซม์ออกไปสลายโปรตีนนอกเซลล์ Autophagic เป็นการสลาย organelle ภายในเซลล์ Phagocytosis เป็นการสลายโปรตีนที่เข้ามาในเซลล์ Autolysis of cell เป็นการสลายตัวเองของเซลล์

46
Lysosome degradation

47
Posttranslational processing
โปรตีนบางชนิดหลังจากสังเคราะห์เสร็จแล้ว จะมีการ เปลี่ยนแปลงภายในโมเลกุลของมันก่อน จึงจะทำงานได้ตามปกติ ขบวนการเปลี่ยนแปลงนี้ ได้แก่ proteolytic cleavage : การเปลี่ยน zymogen หรือ prohormone ให้ เป็น active form covalent modification : γ-carboxylation, hydroxylation, N- methylation, phosphorylation, acylation และ prenylation

48
Proteolytic cleavage การเปลี่ยน zymogen เป็น enzyme ที่ออกฤทธิ์ได้
trypsinogen trypsin factor XII factor XIIa การเปลี่ยน prohormone เป็น hormone proinsulin insulin pro-opiomelanocortin ACTH + MSH + (POMC) β-endorphin
 β-endorphin.")
49
γ-carboxylation เกิดที่ glutamate บางตัวของโปรตีนที่เกี่ยวข้องกับการแข็งตัวของเลือด ปฏิกิริยานี้ต้องอาศัย vitamin K

50
Hydroxylation การเติม –OH ที่ side chain ของกรดอะมิโนบางตัว
ตัวอย่างเช่น hydroxylation ของ proline และ lysine ใน collagen โดยเอนไซม์ proline hydroxylase และ lysyl hydroxylase ได้เป็น hydroxyproline และ hydroxylysine ตามลำดับ ขบวนการนี้จำเป็นต่อการเกิด H-bond ระหว่างสาย collagen ปฏิกิริยานี้ต้องอาศัย vitamin C

51
Phosphorylation การเติมหมู่ phosphate จาก ATP โดยใช้เอนไซม์ protein kinase ATP + protein phosphoprotein + ADP ขบวนการนี้เป็นวิธีที่เซลล์ใช้ควบคุม activity ของเอนไซม์และ โปรตีน กรดอะมิโนที่ถูกเติมหมู่ phosphate ได้ คือ serine, threonine และ tyrosine

52
Glycosylation เป็นการเติมน้ำตาลที่ side chain ของกรดอะมิโน โดยเอนไซม์ glucosyltransferase เกิดขึ้นใน ER และ Golgi body แบ่งเป็น 2 ชนิด N-linked glycosylation O-linked glycosylation การเติมน้ำตาลทำให้โปรตีนจับกับ receptor ที่จำเพาะได้ ( cell recognition) ทำให้โปรตีนมี solubility สูงขึ้น ป้องกันไม่ให้โปรตีนถูกสลายได้ง่าย
 ทำให้โปรตีนมี solubility สูงขึ้น. ป้องกันไม่ให้โปรตีนถูกสลายได้ง่าย.")
53
เอนไซม์ที่ใช้ใน lysosome จะมี mannose-6-phosphate จับที่ส่วนปลาย
Glycosylation เอนไซม์ที่ใช้ใน lysosome จะมี mannose-6-phosphate จับที่ส่วนปลาย ทำให้เอนไซม์จับกับ receptor และเข้า lysosome ได้ ถ้าไม่มี mannose-6-phosphate เอนไซม์จะถูกส่งออกนอกเซลล์ lysosome จึงไม่มีเอนไซม์ เกิดการคั่งของสารใน lysosome (inclusion body) เรียก I-cell disease (mucolipidosis II) ซึ่งเป็น lysosomal storage disease ชนิดหนึ่ง
 เรียก I-cell disease (mucolipidosis II) ซึ่งเป็น lysosomal storage disease ชนิดหนึ่ง.")
54
N-linked glycosylation
เป็นการเติม dolichol phosphate บน amide group ของ asparagine เกิดที่ membrane ของ ER มีผลต่อการเกิด protein folding ถูกยับยั้งด้วย bacitracin และ tunicamycin

55
N-linked glycosylation
High mannose complex hybrid

56
O-linked glycosylation
เป็นการเติมน้ำตาลเข้ากับ –OH group ของ serine หรือ threonine เป็น post-translational process โดยเกิดขึ้นหลังจากที่โปรตีนไปถึง Golgi body ตัวอย่างเช่น ABO blood group antigen H antigen A antigen B antigen

57
Prenylation เป็นการเติม lipid ให้แก่โปรตีน ทำ ให้โปรตีนสามารถ จับกับ membrane ได้ โดยการเติม farnesyl group (C-15) หรือ geranylgeranyl group (C-20) บน cysteine ทางด้าน C-terminal ของ Ras protein
 หรือ geranylgeranyl group (C-20) บน cysteine ทางด้าน C-terminal ของ Ras protein.")
58
Collagen synthesis

59
Collagen โปรตีนที่มีปริมาณมากที่สุดในสัตว์เลี้ยงลูกด้วยนม
โครงสร้างหลักของเนื้อเยื่อเกี่ยวพันหลายชนิด เช่น ผิวหนัง, กระดูก ชนิดของ collagen Type I : 90% ของ collagen ในร่างกาย Type II : พบมากในกระดูกอ่อน Type III : พบมากในหลอดเลือด Type IV : ส่วนประกอบสำคัญของ basement membrane

60
Collagen synthesis ยีน collagen ในมนุษย์ มีมากกว่า 30 ยีน
การเรียกชื่อยีน COL1A2 : ยีนที่ code 2-chain ที่เป็นส่วนประกอบของ type I collagen COL2A1 : ยีนที่ code 1-chain ที่เป็นส่วนประกอบของ type II collagen การเรียกชื่อสาย collagen 1(I) : 1-chain ที่เป็นส่วนประกอบของ type I collagen 1(III) : 1-chain ที่เป็นส่วนประกอบของ type III collagen
 : 1-chain ที่เป็นส่วนประกอบของ type I collagen. 1(III) : 1-chain ที่เป็นส่วนประกอบของ type III collagen.")
61
Collagen synthesis Collagen fibril ประกอบด้วย monomer คือ tropocollagen หลายโมเลกุลที่เชื่อมกันโดยการเกิด cross-link Tropocollagen แต่ละโมเลกุลประกอบด้วย -chain 3 สาย พัน กันเป็น triple helix ชนิดของ -chain ใน collagen type ต่างๆ Type I : 1(I)-chain 2 สาย และ 2(I)-chain 1 สาย Type II : 1(II)-chain 3 สาย (homotrimer) Type III : 1(III)-chain 3 สาย (homotrimer)
-chain 2 สาย และ 2(I)-chain 1 สาย. Type II : 1(II)-chain 3 สาย (homotrimer) Type III : 1(III)-chain 3 สาย (homotrimer)")
62
Biosynthesis of collagen
Nuclear event Transcription : ได้ primary transcript (hnRNA) RNA processing : ได้แก่การเกิด 5’ capping, 3’ polyadenylation และ splicing ได้เป็น mRNA Translation เกิดบน membrand-bound ribosome ได้ prepro -chain ที่มี signal sequence อยู่ทางด้าน N-terminus เพื่อใช้ในการเกิด cotranslational translocation
 RNA processing : ได้แก่การเกิด 5’ capping, 3’ polyadenylation และ splicing ได้เป็น mRNA. Translation. เกิดบน membrand-bound ribosome ได้ prepro -chain ที่มี signal sequence อยู่ทางด้าน N-terminus เพื่อใช้ในการเกิด cotranslational translocation.")
63
Biosynthesis of collagen
Posttranslational modification Signal sequence จะถูกตัดออกจาก prepro -chain กลายเป็น pro -chain Hydroxylation of prolyl and lysyl residues : เกิดเป็น hydroxy - proline (สำคัญต่อ thermal stability) และ hydroxylysine (สำคัญต่อ tissue tensile strength) ขั้นตอนนี้ใช้ vitamin C (ascorbate) เป็น coenzyme O-linked glycosylation : เป็นการเติม galactose หรือ glucose ใน hydroxy lysine N-linked glycosylation : เป็นการเติม mannose-rich CHO ที่ C-terminal ของ pro -chain
 และ hydroxylysine (สำคัญต่อ tissue tensile strength) ขั้นตอนนี้ใช้ vitamin C (ascorbate) เป็น coenzyme. O-linked glycosylation : เป็นการเติม galactose หรือ glucose ใน hydroxy lysine. N-linked glycosylation : เป็นการเติม mannose-rich CHO ที่ C-terminal ของ pro -chain.")
64
Collagen synthesis hydroxylation glycosylation

65
Collagen synthesis

66
Biosynthesis of collagen
Posttranslational modification Procollagen triple helix formation : เกิดการรวมกันของ pro -chain และพันกันเป็นเกลียว ทิศทางการเกิดเริ่มจาก C-terminus ไป N- terminus เรียก triple helix molecule ที่ได้นี้ว่า procollagen Amino-terminal propeptide Carboxyl-terminal propeptide Telopeptide

67
Biosynthesis of collagen
Procollagen = triple helical domain + nonhelical part Nonhelical part = amino-terminal propeptide + telopeptide + carboxyl-terminal propeptide

68
Biosynthesis of collagen
Posttranslational modification Modification beyond RER : procollagen ถูกส่งไปยัง Golgi body และ ส่งออกจากเซลล์ในรูปของ secretory vesicle Extracellular events amino-terminal propeptide และ carboxyl terminal propeptide จะถูกตัดออกโดย procollagen N-protease และ procollagen C- protease ตามลำดับ เรียกโมเลกุลที่ได้ว่า tropocollagen ซึ่งมี ความสามารถในการละลายลดลง 100 เท่า

69
Biosynthesis of collagen
Extracellular events Tropocollagen จะจัดเรียงตัวขนานกันและเหลื่อมกัน 1 ใน 4 ของ ความยาวของโมเลกุล เกิด cross-link เชื่อมระหว่าง tropocollagen โดยเอนไซม์ lysyl oxidase

70
Mutation of collagen genes
Dominant negative effect : การที่มี tropocollagen ที่เกิด จาก mutant collagen gene เข้ามารวมในโมเลกุล collagen fibril ทำให้ collagen fibril นั้นเกิดความผิดปกติไปด้วย Procollagen suicide : abnormal pro -chain (จาก mutant allele) รวมกับ normal pro -chain เกิดเป็น product ที่ถูก สลายไปทั้งหมดก่อนส่งออกนอกเซลล์
 รวมกับ normal pro -chain เกิดเป็น product ที่ถูก สลายไปทั้งหมดก่อนส่งออกนอกเซลล์")
71
Diseases due to abnormal collagen
Osteogenesis imperfecta (brittle bone disease) Bone fragility Mutation ของ type I collagen ส่วนใหญ่มีความผิดปกติของ gene COL1A1 หรือ COL1A2 Type I : mutation ที่ขัดขวางการ splicing และการสร้าง proprocollagen mRNA ทำให้ร่างกายสังเคราะห์ collagen ปกติ ลดลง Type II, III และ IV : point mutation ของ glycine codon ซึ่งจะ ยับยั้งการเกิด normal helix
 Bone fragility. Mutation ของ type I collagen ส่วนใหญ่มีความผิดปกติของ gene COL1A1 หรือ COL1A2. Type I : mutation ที่ขัดขวางการ splicing และการสร้าง proprocollagen mRNA ทำให้ร่างกายสังเคราะห์ collagen ปกติ ลดลง. Type II, III และ IV : point mutation ของ glycine codon ซึ่งจะ ยับยั้งการเกิด normal helix.")
72
Diseases due to abnormal collagen
Osteogenesis imperfecta

73
Diseases due to abnormal collagen
Ehler-Danlos syndrome Skin fragility, skin hyperextensibility และ joint hypermobility Type VI : ความผิดปกติของการเกิด hydroxylation ของ lysine Type VII : ความปกติในการตัด amino-terminal propeptide ออก Type IX : ความผิดปกติในการเกิด cross-linked

74
Diseases due to abnormal collagen
Lathyrism Skin fragility, skeletal deformities และ severe blood vessel weakness สารพิษจากเมล็ดของ sweet pea มีฤทธิ์ยับยั้งเอนไซม์ (lysyl oxidase) ที่ควบคุมการเกิด cross-linked แบบ irreversible Scurvy Vitamin C deficiency Hydroxylation เกิดได้ไม่ดี จึงมีปริมาณ hydroxylysine และ hydroxyproline ใน pro -chain ลดลง ทำให้ collagen stability และ tensile strength น้อยลง
 ที่ควบคุมการเกิด cross-linked แบบ irreversible. Scurvy. Vitamin C deficiency. Hydroxylation เกิดได้ไม่ดี จึงมีปริมาณ hydroxylysine และ hydroxyproline ใน pro -chain ลดลง ทำให้ collagen stability และ tensile strength น้อยลง.")
75
ANY QUESTIONS? THE END

งานนำเสนอที่คล้ายกัน







 ปี 57 เกิด 3 จับ 2 ราย (66.67.00% ) คดีเท่ากัน ผลการจับกุมบรรลุเป้า ( เป้า 75.47 %)>")




>")






