ดาวน์โหลดงานนำเสนอ
งานนำเสนอกำลังจะดาวน์โหลด โปรดรอ

1
บทที่ 9 Photomorphogenesis รองศาสตราจารย์ ดร.ดนัย บุณยเกียรติ
รองศาสตราจารย์ ดร.ดนัย บุณยเกียรติ ภาควิชาพืชสวน คณะเกษตรศาสตร์ มหาวิทยาลัยเชียงใหม่

2
2 1. Phototropism 2. Photoperiodism 3. Photomorphogenesis

3
1. ไฟโตโครม (Phytochrome) 2. คริพโตโครม (Cryptochoe)
3 1. ไฟโตโครม (Phytochrome) 2. คริพโตโครม (Cryptochoe) 3. UV-B photoreceptor 4. โปรโตคลอโรฟิลด์ เอ (Protochlorophyllide a)
 2. คริพโตโครม (Cryptochoe) 3. UV-B photoreceptor. 4. โปรโตคลอโรฟิลด์ เอ (Protochlorophyllide a)")
4
4 Photomorphogenesis มี 2 ระยะ คือ Pattern specification เป็นระยะที่เซลล์และเนื้อเยื่อพัฒนาให้มีความสามารถที่จะตอบสนองต่อแสง และ Pattern realization เป็นระยะที่เป็นกระบวนการที่ตอบสนองต่อแสง ไฟโตโครม เป็นระยะของสารสี ซึ่งพบทั่วไปในพืชชั้นสูงและสาหร่ายมีมวลประมาณ 120,000 ดัลตัน พบในปริมาณที่น้อยมากประมาณ 0.1 % ของโปรตีน ทั้งหมดประกอบด้วยโปรตีนที่มีกลุ่มพรอสทิติค (Prosthetic Group) 1 กลุ่ม ซึ่งเป็นเตตราไพโรลที่เป็นลูกโซ่เปิด (Open-chain tetrapyrrole type) ซึ่งเป็นส่วนที่ดูดแสงได้
 1 กลุ่ม ซึ่งเป็นเตตราไพโรลที่เป็นลูกโซ่เปิด (Open-chain tetrapyrrole type) ซึ่งเป็นส่วนที่ดูดแสงได้")
7
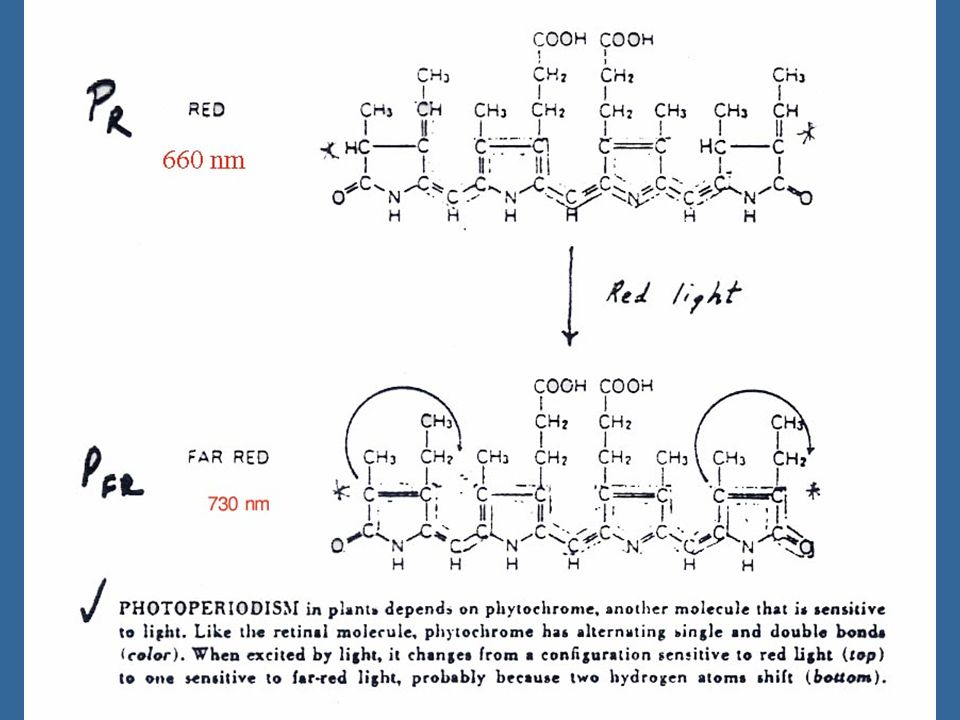
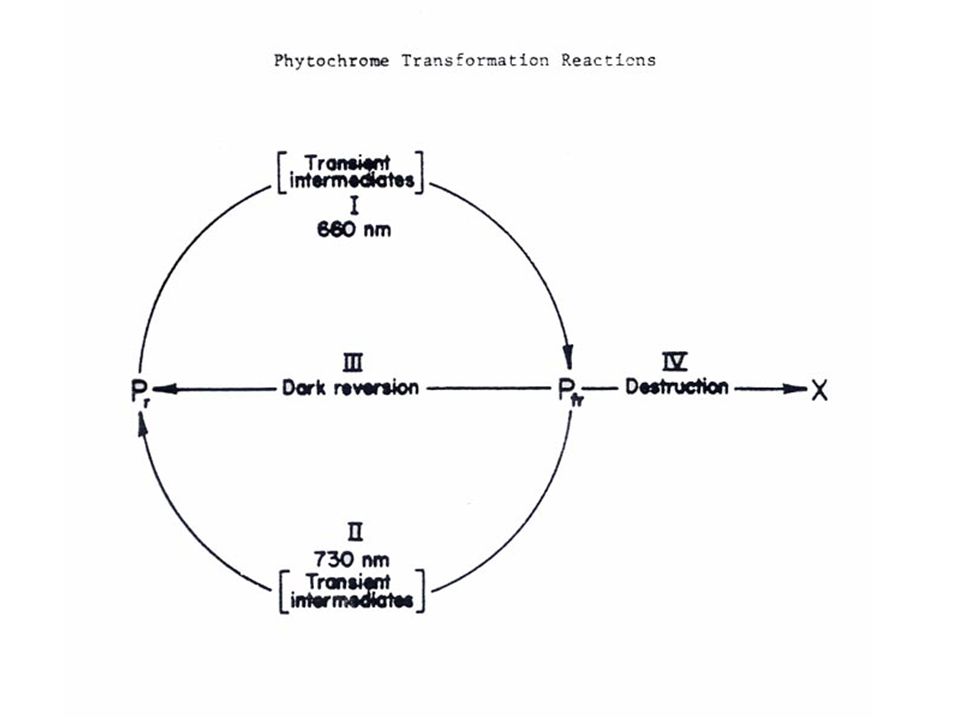
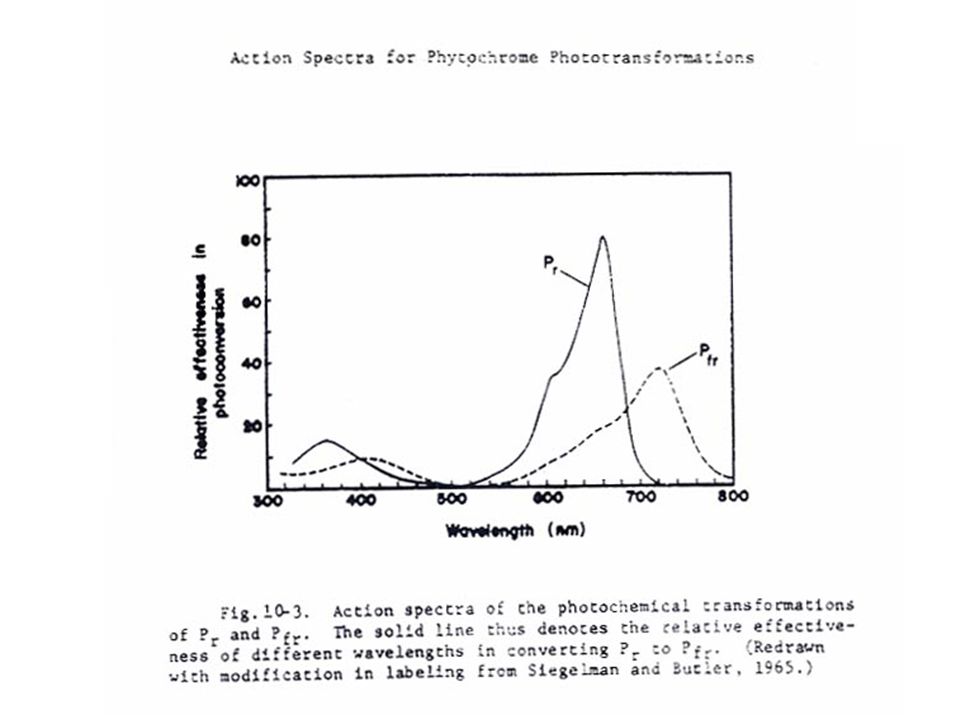
5 ไฟโตโครมที่สามารถดูดแสงสีแดงซึ่งมีความยาวคลื่น nm อยู่ในรูป Pr ซึ่งเป็นรูปที่สังเคราะห์โดยพืช เมื่อได้รับแสงสีแดง Pr จะเปลี่ยนอย่างรวดเร็วเป็น Pfr ซึ่งจะดูดซับแสง Far Red ซึ่งมีความยาวคลื่น nm รูป Pr ของไฟโตโครมเป็นรูปที่ค่อนข้างอยู่ตัว ส่วน Pfr นั้นสลายตัวง่ายในพืชใบเลี้ยงคู่บางชนิด เช่น กะหล่ำดอก Pfr จะสามารถเปลี่ยนกลับไปเป็น Pr ได้ในขณะที่อยู่ในที่มืดเรียกว่า เกิด "Dark Reversion"

10
6 Pfr นั้นเมื่อได้รับแสง Far Red จะเปลี่ยนกลับเป็น Pr อย่างรวดเร็วภายในเวลา มิลลิเซคกัน (milliseconds) ส่วน Pr จะกลับเป็น Pfr เมื่อได้รับแสงสีแดงเป็นเวลาหลายวินาที โดยทั่วไปแสงสีแดงจะทำให้เกิด Pfr ประมาณ 80 เปอร์เซ็นต์ และ Pr 20 เปอร์เซ็นต์
 ส่วน Pr จะกลับเป็น Pfr เมื่อได้รับแสงสีแดงเป็นเวลาหลายวินาที โดยทั่วไปแสงสีแดงจะทำให้เกิด Pfr ประมาณ 80 เปอร์เซ็นต์ และ Pr 20 เปอร์เซ็นต์")
11
สาหร่าย Bryophytes และ Pteridophytes
7 สาหร่าย Bryophytes และ Pteridophytes การงอกของสปอร์ การเคลื่อนที่ของคลอโรพลาสต์ การเจริญและพัฒนาของ Protonema

12
Gymnosperms การงอกของเมล็ด การโค้งของส่วนใต้ใบเลี้ยงของต้นอ่อน
8 Gymnosperms การงอกของเมล็ด การโค้งของส่วนใต้ใบเลี้ยงของต้นอ่อน การขยายตัวของปล้อง การพักตัวของตา

13
Angiosperms การงอกของเมล็ด การโค้งของส่วนใต้ใบเลี้ยงของต้นอ่อน
9 Angiosperms การงอกของเมล็ด การโค้งของส่วนใต้ใบเลี้ยงของต้นอ่อน การขยายตัวของปล้อง การเกิดส่วนที่เจริญเป็นราก การเกิดใบและการเจริญเติบโต การเคลื่อนที่ของใบย่อย การยอมให้สารผ่านเข้าออกของเยื่อหุ้มเซลล์ การโค้งเข้าหาและออกจากแสง การตอบสนองต่อแรงดึงดูดของโลก การสังเคราะห์แอนโธไซยานิน

14
10 ไฟโตโครมนั้นมีสองชนิดเรียกว่า ชนิดที่ 1 (Type I) และชนิดที่ 2 (Type II) ชนิดที่ 1 นั้น พบมากในต้นอ่อนที่เจริญเติบโตในที่มืด (Etiolated seeding) และชนิดที่ 2 พบมากในพืชสีเขียว และเมล็ด (อย่างน้อยพบในเมล็ดของข้าวโอ๊ต) จากการศึกษาปฏิกิริยาทาง Immunology โปรตีนของไฟโตโครมชนิดที่ 1 และ 2 ต่างกันอย่างชัดเจน
 และชนิดที่ 2 (Type II) ชนิดที่ 1 นั้น พบมากในต้นอ่อนที่เจริญเติบโตในที่มืด (Etiolated seeding) และชนิดที่ 2 พบมากในพืชสีเขียว และเมล็ด (อย่างน้อยพบในเมล็ดของข้าวโอ๊ต) จากการศึกษาปฏิกิริยาทาง Immunology โปรตีนของไฟโตโครมชนิดที่ 1 และ 2 ต่างกันอย่างชัดเจน.")
15
11 การสลายตัวของ Pfr ของไฟโตโครมชนิดที่ 1 นั้น ในปัจจุบันทราบแน่ชัดแล้วว่าเกิดโดยกระบวนการ ซึ่ง Pfr จะไปเกาะติดกับโปรตีนขนาดเล็ก ซึ่งชื่อว่า Ubiquitin ซึ่งพบในเซลล์ของสิ่งมีชีวิตชนิด Eucaryote ทั่วไป แต่ไม่พบใน ProcaryoteUbiq uitin เป็นโปรตีนขนาดเล็ก ซึ่งมีกรดอะมิโนเพียง 76 ลำดับ การเกาะติดกับ Ubiquitin ของ Pfr มีผลทำให้เกิดการทำลาย Pfr โดยการคะตะไลท์ของเอนไซม์ 3 ชนิด และเป็นกระบวนการซึ่งต้องการ ATP

16
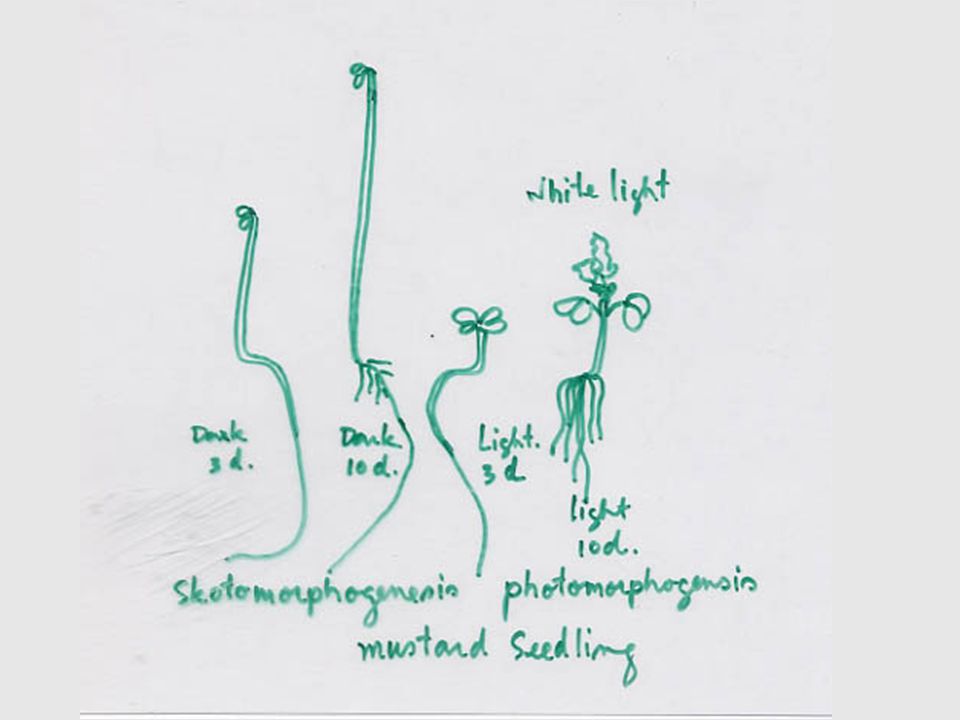
Skotomorphogenesis Photomorphogenesis
12 Skotomorphogenesis Photomorphogenesis 1. ลักษณะทางสัณฐานวิทยา - ใบเลี้ยงไม่คลี่ขยายมีขนาดเล็ก -มี Hook บริเวณส่วนใต้ใบเลี้ยง - ส่วนใต้ใบเลี้ยงยาวมาก - ใบเลี้ยงคลี่ขยายแผ่ใบเพื่อการรับแสง Hook จะหายไปทำให้ต้นตั้งตรง ส่วนใต้ใบเลี้ยงมีลักษณะปกติ 2. เซลล์ของใบเลี้ยง (Cotyledon) - สร้าง Ethioplast -ไม่มีปากใบหรือมีน้อยมาก - สร้าง Glyoxysome ซึ่งย่อยสลายไขมัน - สร้าง Chloroplast ซึ่งมีเม็ดแป้งภายใน - มีปากใบตามปกติ - สร้าง Peroxisome 3. ลักษณะทางสรีรวิทยาของใบเลี้ยง -ไม่สะสมแอนโธไซยานิน -หายใจและเคลื่อนย้ายสารอินทรีย์เร็ว - สะสมแอนโธไซยานิน - หายใจปกติและเคลื่อนย้ายสารอินทรีย์ช้า
 - สร้าง Ethioplast. -ไม่มีปากใบหรือมีน้อยมาก. - สร้าง Glyoxysome ซึ่งย่อยสลายไขมัน. - สร้าง Chloroplast ซึ่งมีเม็ดแป้งภายใน. - มีปากใบตามปกติ - สร้าง Peroxisome. 3. ลักษณะทางสรีรวิทยาของใบเลี้ยง. -ไม่สะสมแอนโธไซยานิน. -หายใจและเคลื่อนย้ายสารอินทรีย์เร็ว. - สะสมแอนโธไซยานิน. - หายใจปกติและเคลื่อนย้ายสารอินทรีย์ช้า.")
19
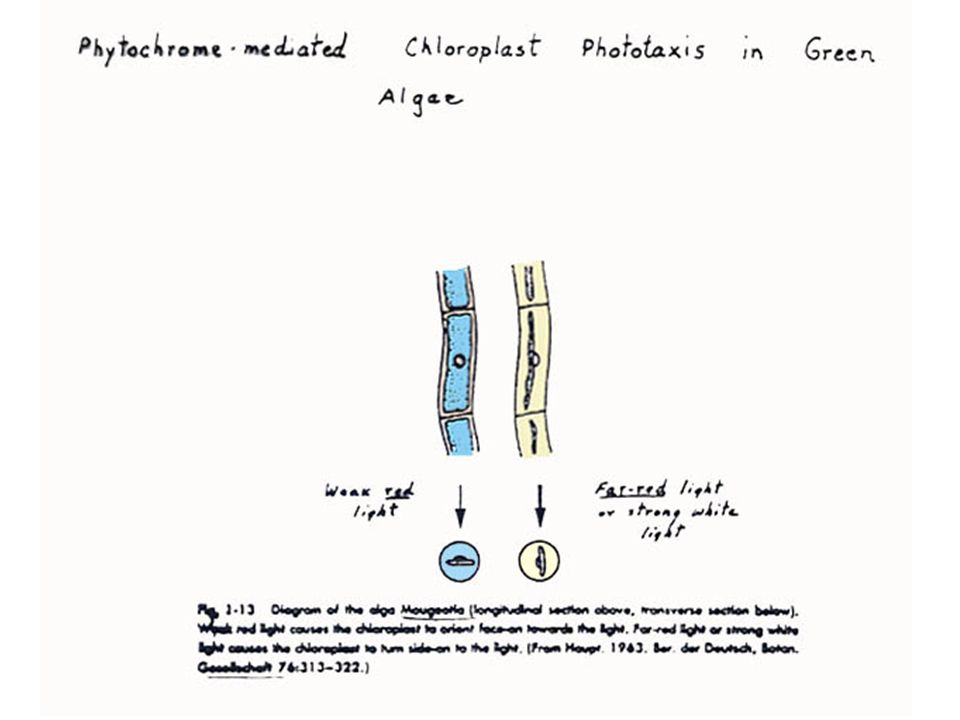
ผลของแสงต่อการเรียงตัวของคลอโรพลาสต์13
เมื่อความเข้มของแสงสูง ตามปกติคลอโรพลาสต์จะเรียงตัวในแนวตรงไปตามรัศมีของผนังเซลล์ เพื่อให้บังร่มกันเองป้องกันการถูกทำลายจากแสง ในสภาพแสงความเข้มน้อยหรือในที่มืดคลอโรพลาสต์จะแยกออกเป็นสองกลุ่มกระจายไปตามผนังเซลล์ที่ใกล้ที่สุดจากแหล่งแสงเพื่อให้ดูดรับแสงให้ได้มากที่สุด

21
ผลของแสงในการกระตุ้น การสังเคราะห์ฟลาโวนอยส์ (Flavonoid)
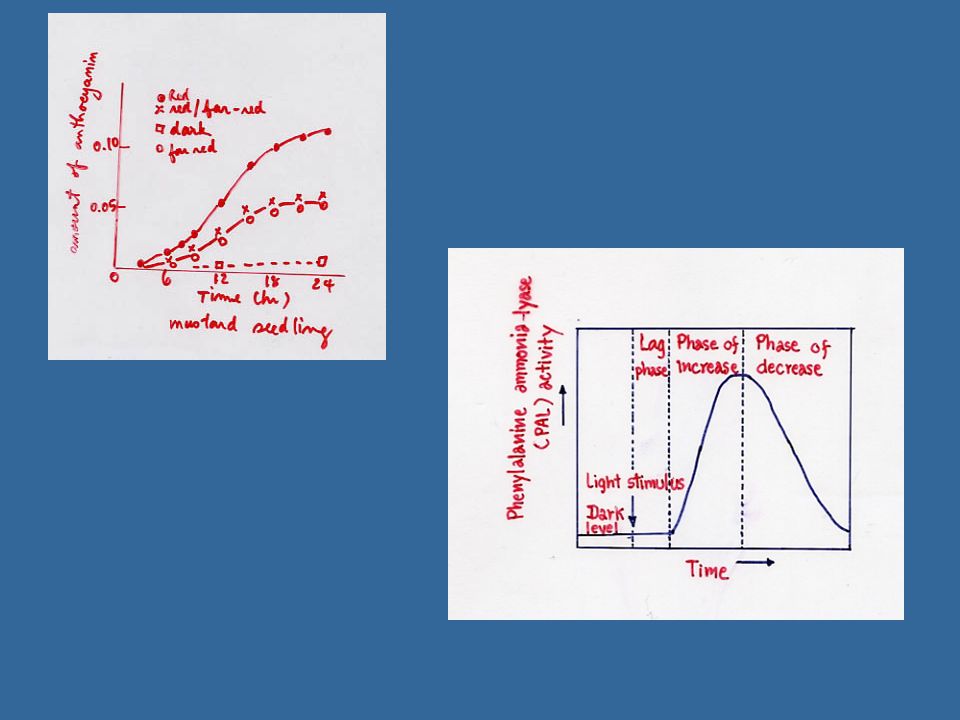
14 ผลของแสงในการกระตุ้น การสังเคราะห์ฟลาโวนอยส์ (Flavonoid) ผลแอปเปิลที่อยู่ทางด้านใต้ทรงพุ่มจะสังเคราะห์ แอนโธไซยานินได้มากกว่าและเร็วกว่าผลที่อยู่ทางด้านเหนือของทรงพุ่ม การสังเคราะห์ฟลาโวนอยส์ต้องการน้ำตาลเพื่อเกิดเป็น Phosphoenolpyruvate และ Erythrose-4-phosphate ซึ่งเป็นแหล่งของคาร์บอนในวงแหวน B และเป็นแหล่งของกลุ่มอะซีเตทซึ่งอยู่ในวงแหวน A
 ผลแอปเปิลที่อยู่ทางด้านใต้ทรงพุ่มจะสังเคราะห์ แอนโธไซยานินได้มากกว่าและเร็วกว่าผลที่อยู่ทางด้านเหนือของทรงพุ่ม. การสังเคราะห์ฟลาโวนอยส์ต้องการน้ำตาลเพื่อเกิดเป็น Phosphoenolpyruvate และ Erythrose-4-phosphate ซึ่งเป็นแหล่งของคาร์บอนในวงแหวน B และเป็นแหล่งของกลุ่มอะซีเตทซึ่งอยู่ในวงแหวน A.")
22
ผลของแสงในการกระตุ้น การสังเคราะห์ฟลาโวนอยส์ (Flavonoid) 15
การใช้ Phenylalanine ในการสร้างวงแหวน B เพราะว่า Phenylalanine นั้นสามารถถูกนำไปใช้ในกระบวนการเมตาบอลิสม์เป็นจำนวนมาก การควบคุมของแสงเป็นขั้นตอนแรกในการเปลี่ยน Phenylalanine ให้เกิดวงแหวน B ขั้นตอนนี้ต้องการเอนไซม์ชื่อ Phenylalanine ammonia lyase และแสงจะกระตุ้นกิจกรรมของเอ็นไซม์นี้ในพืชหลายชนิด ในทำนองเดียวกันลิกนินซึ่งเกิดโดย Shikimic pathway เหมือนฟลาโวนอยด์ก็เกี่ยวข้องกับเอนไซม์ Phenylalanine ammonia lyase

24
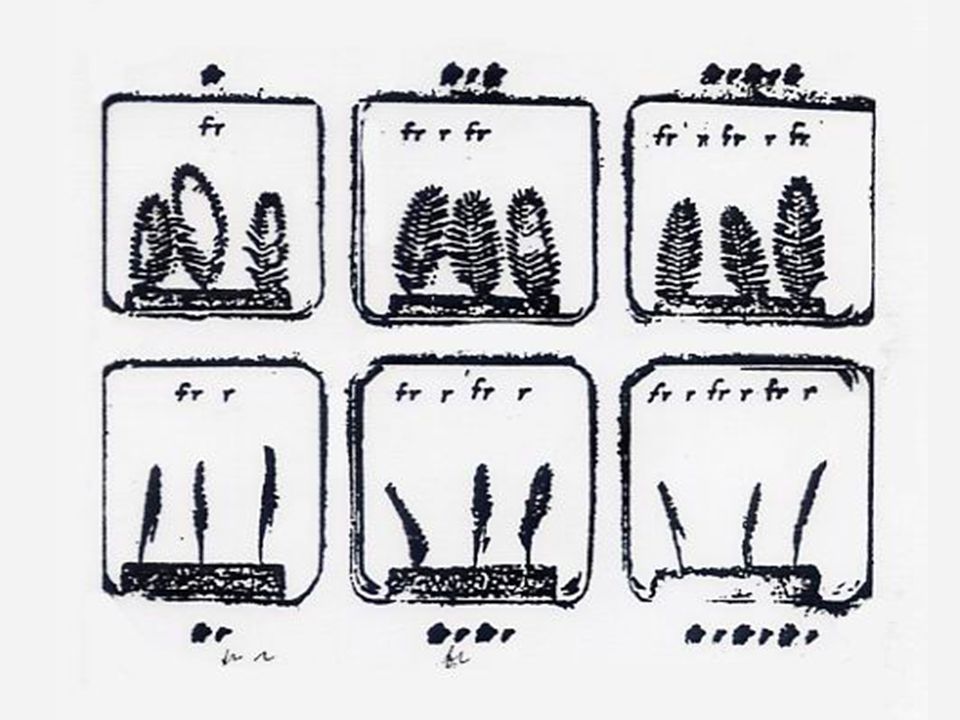
ผลของแสงในการงอกของเมล็ด
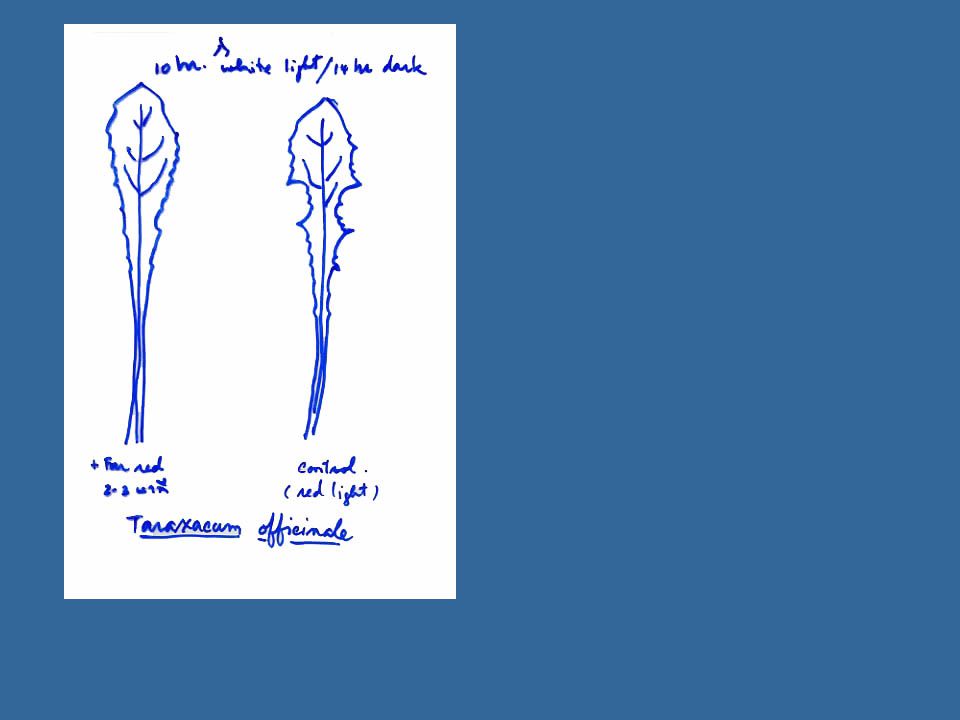
16 ผลของแสงในการงอกของเมล็ด เมล็ดที่ต้องการแสงในการงอกเรียกว่า Photodormant ระดับอุณหภูมิก่อนและหลังการได้รับแสงจะมีความสำคัญมากกว่าในระหว่างการได้รับแสงโดยตรง กระบวนการทางเคมีซึ่งควบคุมโดย Phytochrome จะมีความไวต่ออุณหภูมิ เมล็ดของผักสลัดพันธุ์ Grand Rapids ซึ่งปกติแสงจะกระตุ้นการงอกแต่ถ้าให้เมล็ดอยู่ที่อุณหภูมิ 35 oC หลังจากการได้รับแสงหรือในขณะได้รับแสงจะทำให้เมล็ดนี้พักตัว เมล็ดของผักสลัดพันธุ์ Great Lakes ไม่ต้องการแสงในการงอก แต่ถ้าหากเมล็ดที่ชื้นอยู่ภายใต้อุณหภูมิ 35 oC เมล็ดนี้จะกลายเป็นเมล็ดชนิด Photodormant

25
LIGHT TREATMENT GERMINATION Dark conrol 8.8 Red (1 min) 98 Red : Far-red (1 min) 54 R : F-R : R 100 R : F-R : R : F-R 43 R : F-R : R : F-R :R 99

28
กลไกการทำงานของไฟโตโครม
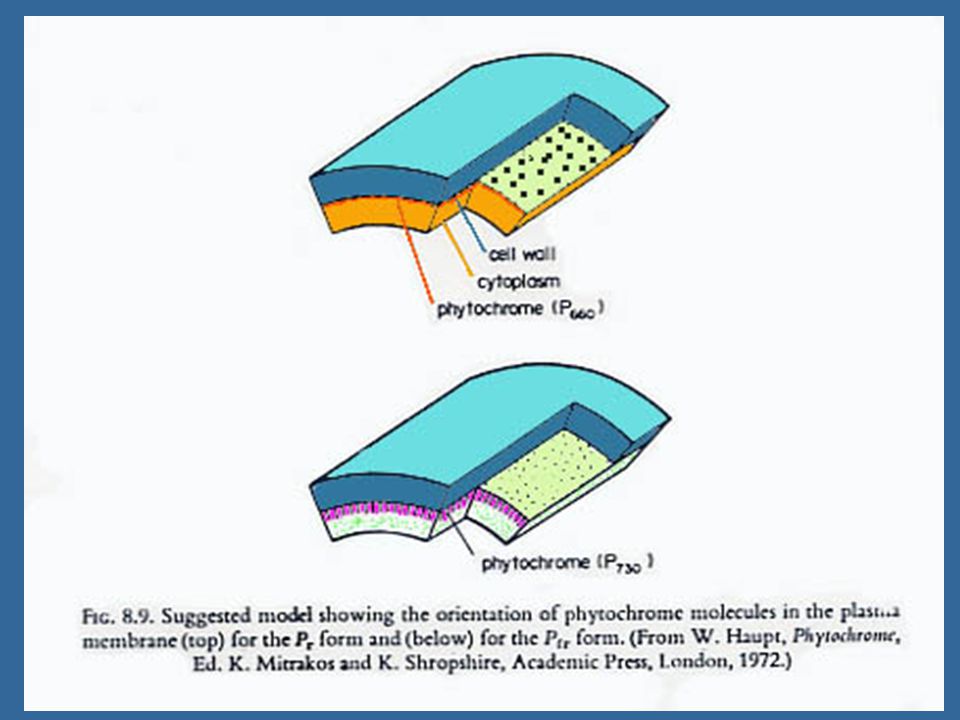
17 กลไกการทำงานของไฟโตโครม การเปลี่ยนแปลงการยอมให้สารผ่านเข้าออกของเยื่อหุ้มเซลล์ (Change in Membrane Permeability) ผลในขั้นทีสองของไฟโตโครม คือ การควบคุมระบบเอนไซม์
 ผลในขั้นทีสองของไฟโตโครม คือ การควบคุมระบบเอนไซม์")
งานนำเสนอที่คล้ายกัน





และฝน “เมฆ” อากาศเคลื่อนตัวสูงขึ้นถึงระดับที่อากาศมีอุณหภูมิ ต่ำ จนอากาศอิ่มตัวด้วยไอน้ำ ไอน้ำจะเกิดการควบแน่นเป็น ละอองน้ำหรือระเหิดกลับเป็นผลึกน้ำแข็งขนาดเล็ก.>")







.>")
.>")





>")